驯化xnmsn.cn
xnmsn.cn 时间:2021-04-07 阅读:()
基金项目国家自然科学基金项目(No.
30560026),云南科技强省重点项目(No.
2007C000Z1);*通讯作者,Email:wzk-930@yahoo.
com.
cn;第一作者介绍张麟,男,大学本科生;研究方向:动物生理生态学;Email:lzhangss@xnmsn.
cn.
收稿日期:20081229,修回日期:20090513动物学杂志ChineseJournalofZoology2009,44(4):47~57中缅树肝在冷适应条件下的产热特征张麟王睿杨芳谢静王政昆*龚正达郭宪国!
(云南师范大学生命科学学院教育部生物能源持续开发利用工程研究中心云南省能源与环境生物技术重点实验室昆明650092;大理学院基础医学部媒介与病原生物研究所云南大理671000;!
云南流行病防治研究所云南大理671000)摘要:中缅树(Tupaiabelangeri)为东洋界特有小型哺乳动物.
本研究测定了中缅树冷驯化实验组(7d、14d、21d、28d)与对照组(0d)中肝线粒体蛋白含量、呼吸状态和状态#的变化,探讨中缅树对不同冷环境的适应情况以及肝产热的机理.
结果表明,在冷驯化条件下,中缅树产热显著增加,与对照组相比,实验组肝总蛋白含量、肝线粒体蛋白含量、呼吸状态和状态#有着显著的提高,在28d后分别增加了399%、393%、849%、1811%,因此,肝在中缅树冷适应产热过程中具有重要的作用.
同时,从生理生态方面为树的岛屿起源学说提供依据.
关键词:中缅树;冷适应;肝;产热特征;岛屿起源中图分类号:Q958,Q955文献标识码:A文章编号:02503263(2009)044711ThermogenesisCharacteristicsofColdAdaptationofLiverinTupaiabelangeriZHANGLinWANGRuiYANGFangXIEJingWANGZhengKun*GONGZhengDaGUOXianGuo!
(SchoolofLifeScienceofYunnanNormalUniversity,EngineeringResearchCenterofSustainableDevelopmentandUtilizationofBiomassEnergyMinistryofEducation,TheKeyLaboratoryofBiomassEnergyandEnvironmentBiologicalTechnologyinYunnanProvince,Kunming650092;LaboratoryofVectorandPathogenofBasicMedicineFaculty,DaliUniversity,YunnanDali671000;!
YunnanInstituteofEpidemicDiseaseControlandResearch,YunnanDali671000,China)Abstract:TheTreeShrew(Tupaiabelangeri)isauniquespeciesofsmallmammalsintheOrientalrealm.
Inordertostudytheircoldadaptivecapacityincoldenvironment,thestateandthe#stateofmitochondrialrespiration,theproteincontentofmitochondriaofliverweremeasuredundertheconditionofbeingcoldexposure(51%,12L&12D)duringdifferenttimes[0d(control),7d,14d,21d,28d].
Comparedwiththecontrols,thetotalprotein,mitochondrialprotein,andthestateandthe#stateofmitochondrialrespirationoflivergreatlyincreasedwithprolongedcoldexposure(increased399%,393%,849%,1811%,respectively,after28d).
TheresultindicatesthattheliverplaysakeyroleoftheadaptivethermogenesisduringcoldexposureintheT.
belangeri.
ThephysiologicalecologyevidenceisalsogiventotheislandoriginofT.
belangeri.
Keywords:Tupaiabelangeri;Coldadaptation;Liver;Thermogenesischaracteristics;Islandorigin在应对冬季的冷胁迫时,许多冬季活动的小型哺乳动物增加产热来维持稳定的体温[1].
在低温胁迫环境中,小型哺乳动物的生理和生化机制发生变化,这些变化表现出对生活环境的适应[2],并影响到动物的生理机制和进化机制[3],这方面的研究已经成为动物生理生态的重要研究领域.
国内外对小型哺乳动物适应性产热的研究主要集中在能量代谢、体重、体温及激素分泌的季节性调节上,但多数研究对象是典型的北方动物,热带种类研究较少[4~6].
肝是恒温动物尤其是小型动物的主要产热器官[7],动物的基础代谢率中有20%~25%来自肝[8],而冷驯化正是刺激基础代谢率增加的重要因素[9].
在线粒体水平上,个体产热受到冷驯化影响显著,中缅树(Tupaiabelangeri)肝线粒体呼吸也显著受到低温的影响.
中缅树在低温环境中,为了适应冷胁迫,其产热可能与质子漏有关[10].
中缅树属于攀目(Scandentia)树科(Tupaiidae)树属,主要分布于东南亚、印度、锡金、尼泊尔、缅甸及我国广西南部和海南岛等地,为东洋界特有小型哺乳动物,是该科中分布最广和分布纬度最高的一个种[11].
中缅树在我国的分布范围,为青藏高原南延部分,其海拔西部高东部低,相对高度变化较大,属典型的热带、亚热带气候地区,其环境条件由湿热类型向干旱寒冷逐渐转变,形成明显的气候过渡地带,为动物生存提供了多种多样的立体栖息环境;尤其横断山脉地区,为北方耐寒动物种类向南迁移和南方热带种类向北扩散提供了通道[12].
我国的云贵高原可能是中缅树分布的北限[11],分布于该过渡地带的中缅树,其产热能力显示出了温带动物的某些特征,因此其生理适应上可能会显示出与东洋界其他小型哺乳动物的不同.
本实验室已有的实验研究表明,分布于该过渡地带的中缅树,其生理适应特征显示出某些过渡特征,同时又表现出一定的差异性[13].
通过测定中缅树冷驯化实验组(7d、14d、21d、28d)与对照组(0d)中肝线粒体蛋白含量、呼吸状态和状态#的变化,探讨中缅树对不同冷环境的适应情况以及肝产热的机理.
1材料与方法11实验动物实验动物于2006年7月捕自云南省昆明市禄劝县屏山镇(北纬2525(~2622(,东经10213(~10257()附近灌丛中(海拔1679m),动物捕回后单笼饲养,22~25%室温条件下适应3d,光照12L&12D,此为0d对照组.
冷驯化7d、14d、21d、28d共4组,每组8只动物,在环境温度(51)%、无巢材、光照12L&12D条件下单笼饲养.
每天上午定点喂以由玉米面、奶粉、白糖以90%&5%&5%比例混合制得的熟食,添加少许水,每隔一天加喂苹果、梨等水果适量.
实验动物均为非繁殖期成年个体,实验组与对照组中尽量雌雄各半,饲养地点为云南师范大学生命科学学院动物学实验室.
12耗氧量测定121基础代谢率(basalmetabolicrate,BMR)的测定动物的基础代谢率采用澳大利亚生产ADML870开放式呼吸仪测定.
代谢率的计算方法为Hill[14]所述,RMR=F(FIO2-FEO2)M(1-FEO2),式中,F为标准状态下的气体流量(mlmin),FIO2为进入呼吸室气体的氧气浓度,FEO2为流出呼吸室气体的氧气浓度(mlmin),RMR为静止代谢率(mlO2g)h),M为体重(g).
将呼吸室置于上海博迅医疗设备厂生产的SPX300型人工气候箱,由其将实验温度控制在中缅树的热中性区(3005)%[15].
呼吸室为500ml,进入呼吸室的气体流量为200mlmin.
动物实验前一般饥饿2~4h,实验进行15h,进入呼吸室后适应30min,然后记录1h测定基础代谢率.
实验前后均称体重,以实验前后体重(精确到01g)的平均值作为本次的动物体重.
实验前)48)动物学杂志ChineseJournalofZoology44卷后均测体温,采用数字式体温计(北京师范大学司南仪器厂)插入直肠约3cm测量动物的体温,每次测温探头在直肠内静置约45s左右读数,并以实验后体温作为动物在该温度下的体温.
122非颤抖性产热(nonshiveringthermogenesis,NST)的测定在BMR测定后,迅速取出实验动物,在肩胛间注射去甲肾上腺素(norepinephrine,NE),用ADML870开放式呼吸仪测定动物在30%下的最大非颤抖性产热(themaximalnonshiveringthermogenesis,NSTmax).
注射与体重相应剂量10mgkgBM(体重bodymass,BM)的NE(剂量经预备实验确定),再放回呼吸室连续测定30min左右,选取最大持续耗氧量作为NSTmax[16].
13肝组织分离线粒体及线粒体蛋白含量、呼吸活性测定131肝线粒体的分离动物用断颈法处死后,迅速取出肝.
按照Cannon等[17]介绍的方法,将肝在冰浴中剪碎,加5倍体积(wv)肝提取液,手动匀浆;2000g4%离心7min,弃沉淀;取上清于10000g4%离心10min,弃上清;将沉淀用适量提取液悬浮,于10000g4%离心10min,弃上清;所得沉淀即为线粒体,加适量提取液溶解,即可用于测定呼吸活性.
132肝线粒体呼吸活性测定采用CHLOTOLAB2氧电极(HansatechInstrumentsLab)测定线粒体呼吸状态和状态#.
加5molADP测定状态呼吸,ADP耗尽后变为线粒体呼吸状态#;呼吸底物为琥珀酸,反应杯总体积为2ml,反应温度为30%.
133肝线粒体含量测定蛋白质定量采用FolinPhenol法,以牛血清白蛋白为标准.
14数据处理实验数据用SPSS150统计软件包进行分析处理,体重、体温、基础代谢率、非颤抖性产热、肝重量以及肝线粒体呼吸指标采用回归分析,所有测量指标均采用单因子方差(OneWayANOVA)多重比较,并且用协方差分析处理体重对基础代谢率、非颤抖性产热的影响.
结果均以MeanSE表示,P005).
随着冷暴露时间的延长,体重逐渐增加,到21d时,体重变化达到显著水平(P005).
随着冷暴露时间的延长,中缅树的体温与冷驯化时间呈显著负相关(P005).
驯化时间对基础代谢率的影响极显著(F4,35=19084,P005).
驯化时间对最大非颤抖性产热的影响极显著(F4,35=639,P<001),而且随着驯化时间的延长,最大非颤抖性产热会增加.
NSTmax与驯化时间(D)显著相关,两者之间的回归方程为:NSTmax=237+0164D(r2=092,P<0001,n=40)(图2).
在冷驯化过程中,最大非颤抖性产热对中缅树肝线粒体产热的贡献随着冷驯化时间的延长而显著降低,21d达到极显著水平,驯化图2冷驯化对中缅树NSTmax的影响Fig.
2TheeffectsofcoldexposureonNSTmaxinTupaiabelangeri与对照组(0d)比较,**P<001.
**P<001,comparedwithcontrol(0d).
28d后比对照组下降了745%(P<001),与对照组(0d)比较差异极显著(OneWayANOVA,F4,35=4944,P<001),并且与驯化时间(D)显著相关,两者之间的回归方程为:NSTmax-BMRBMR100%=4953-825D(r2=087,P<001,n=40)(图3).
23肝重量(theweightofliver,W)及肝相对重量(therelativeweightofliver,WX)的变化中缅树肝重量及肝的相对重量均随着冷驯化时间的延长而显著增加,14d时即达到极显著水平,分别比对照组增加了193%和168%(P<001).
28d均达到极显著水平,肝重量与对照组(0d)比较差异极显著(OneWayANOVA,F4,35=22117,P<001),同时肝相对)50)动物学杂志ChineseJournalofZoology44卷图3冷驯化中缅树最大非颤抖性产热(NSTmax)对产热的贡献Fig.
3ThecontributionoflowtemperatureonNSTforthermogenesisadaptiveofTupaiabelangeri与对照组(0d)比较,*P<005,**P<001.
*P<005,**P<001,comparedwithcontrol(0d).
重量也出现极显著差异(OneWayANOVA,F4,35=8223,P<001),分别比对照组(0d)增加516%和371%(P<001).
二者均与驯化时间(D)显著相关,两者之间的回归方程分别为:W=384+0584D(r2=098,P<001,n=40);WX=378+0349D(r2=097,P<001,n=40)(图4).
图4冷驯化对中缅树肝重量的影响Fig.
4TheeffectsoflowtemperatureontheweightofliverinTupaiabelangeri与对照组(0d)比较,**P<001.
**P<001,comparedwithcontrol(0d).
24肝组织中总蛋白(theproteincontentofliver,TP)与线粒体蛋白(theproteincontentofmitochondria,MP)含量的变化中缅树肝总蛋白含量随着驯化时间的增加而显著增加(P<001),7d时开始达到显著水平,比对照组(0d)增加了149%(P<005);到28d时增加了399%(P<001),与对照组(0d)相比出现极显著差异(OneWayANOVA,F4,35=53423,P<001).
并且TP与驯化时间(D)显著相关,两者之间的回归方程为:TP=8831+917D(r2=098,P<001,n=40).
肝线粒体蛋白含量也随着驯化时间的增加而显著增加(P<001),在14d时达到显著水平,比对照组增加了140%(P<005);28d时增加了393%(P<001),与对照组(0d)相比出现极显著差异(OneWayANOVA,F4,35=24469,P<001).
并且MP与驯化时间(D)显著相关,两者之间的回归方程为:MP=2743+349D(r2=092,P<001,n=40)(图5).
图5冷驯化对中缅树肝蛋白质含量的影响Fig.
5TheeffectsoflowtemperatureonthecontentofproteininliverofTupaiabelangeri与对照组(0d)比较,*P<005,**P<001.
*P<005,**P<001,comparedwithcontrol(0d).
25肝线粒体状态呼吸(thestateofmitochondrialrespiration,ST)及状态#呼吸(the#stateofmitochondrialrespiration,ST#)的变化中缅树肝线粒体状态呼吸随着冷驯化时间的增加而显著增加(P<001),7d时即达到极显著水平,比对照组增加了208%(P<001);到28d时增加了849%,ST含量与对照组(0d)相比出现极显著差异(OneWayANOVA,F4,35=54732,P<001).
ST与驯化时间(D)显著相关,两者之间的回归方程为:ST=00341+00091D(r2=098,P<001,n=40).
肝线粒体状态#呼吸也随着驯化时间)51)4期张麟等:中缅树肝在冷适应条件下的产热特征的增加而显著增加(P<001),在7d时达到极显著水平,比对照组增加了545%(P<005),28d时增加了1811%,ST#含量与对照组(0d)相比出现极显著差异(OneWayANOVA,F4,35=58037,P<001).
ST#与驯化时间(D)显著相关,两者之间的回归方程为:ST#=00083+00056D(r2=098,P<001,n=40)(图6).
图6冷驯化对中缅树肝线粒体呼吸的影响Fig.
6TheeffectsoflowtemperatureontherespirationofmitochondrialinliverofTupaiabelangeri与对照组(0d)比较,**P<001,**P<001,comparedwithcontrol(0d).
3讨论线粒体是哺乳动物消耗氧气的主要部位[18],肝是氧气消耗的主要器官,承担呼吸率的15%~20%[17],低温对线粒体影响显著,而肝受到低温的影响更为显著[19].
肝在冷胁迫下对中缅树的产热有着不可忽视的作用.
31体重、体温与冷驯化的关系Nagy等认为小型哺乳动物冬季的能量收支有两种相反的进化对策:冬眠哺乳动物在秋季体重(大多为脂肪)急剧增加,冬眠前能量的大量贮存与冬眠时低代谢率使得动物顺利过冬;而非冬眠小型哺乳动物在秋季体重下降,从而降低了总体代谢率和对能量的需要[20].
在冷驯化过程中,中缅树的体重不能维持恒定:随着驯化时间的延长,中缅树的体重呈现逐渐增加的趋势(表1).
但在冷驯化7d时,中缅树的体重下降,这可能是其在新的生活环境下形成了新的生理适应来应对低温对其的影响;21d与对照组(0d)相比差异显著,28d后体重比对照组增加100%.
这种变化趋势与同地区的啮齿类动物高山姬鼠(Apodemuschevrieri)和大绒鼠(Eothenomysmiletus)不同,后两种啮齿类动物在冷驯化28d后体重比对照组(0d)分别减少1605%和1101%[4],而且与黑线毛足鼠(Phodopussungorus)、属(Sorex)的大多数种类等典型北方小型哺乳动物在冬季或冷驯化条件下体重降低[21]的趋势亦不同.
中缅树在冷驯化过程中体重随着驯化时间的延长,呈现逐渐增加的趋势,与环颈旅鼠(Dicrostonyxgroenlandicus)和金色中仓鼠(Mesocricetusauratus)在低温胁迫下,体重的变化相同[22].
在冷驯化过程中,中缅树体温随冷驯化时间的延长而逐渐下降(表1),这与中缅树生活在年温差较小,日温差较大,冬短无夏,春秋季长达9个多月的生活环境中有关,也与之前的研究结果一致[23].
中缅树的体温还具有明显的昼夜变化节律,体温的昼夜变化幅度达到5%以上,这一特征对其抵抗夜间低温也具有重要的适应意义[24].
小型哺乳动物的代谢产热特征和体温调节与其能量利用、分配、生活史对策及其进化途径等方面密切相关,反映了动物对环境的适应模式和生理能力[25].
在低温胁迫下,中缅树可以通过降低体温来调节能量分配,减少与环境温度的差值,以达到减少散热、节省能源[26]的目的;同时,增加体重可减少相对体表面积,降低热传导,进一步增强低温适应能力.
由于中缅树散热能力比产热能力强[15],通过增加体重、降低体温,使其在增加产热相对不多的条件下,减少散热、用于体温调节的能量,这对其生存具有重要的适应意义.
32BMR与NSTmax的变化关系基础代谢率BMR是生理生态学中的重要指标,能反映不同物种、个体之间的能量消耗水平,在适应环境的过程中具有重要的意义[22],它受到很多因素如温度、食物等影响[27],也与季节性变化有关[28].
)52)动物学杂志ChineseJournalofZoology44卷增强产热能力是小型哺乳动物抵抗低温胁迫的一种重要生理适应对策[29].
在冷胁迫环境中,小型哺乳动物可以通过提高BMR和NSTmax来增强冷适应能力[30].
大量研究结果表明,低温是增加小型哺乳动物产热的主要因子:黑田鼠(Microtusagrestis)在10%冷驯化100d后,BMR增加522%[31];小家鼠(Musmusculus)在持续降低环境温度的情况下,BMR持续上升,实验组(5%)BMR比对照组增加23%[32];我们的结果,中缅树冷驯化后BMR显著增加,实验组(5%)BMR比对照组增加5543%,也表明低温是刺激中缅树BMR增加的主要因子.
但是与典型北方小型哺乳动物不同的是,低温刺激中缅树BMR增加的程度较高.
黑田鼠、小家鼠等均为典型北方小型哺乳动物,在低温胁迫条件下,其BMR绝对值比中缅树对照组(未冷驯化)高,但比接受冷驯化的中缅树BMR增加程度低;但中缅树接受冷驯化后BMR增加程度比同地区的大绒鼠(737%)低[33],这与中缅树产热比散热少相一致.
低温对小型哺乳动物BMR的影响与动物的地理分布有关,所以BMR在增强中缅树抵抗低温胁迫能力中起着非常重要的作用,同时也说明低温可能是影响中缅树进一步向北扩展的限制因素之一.
冷胁迫情况下,NSTmax在提高小型哺乳动物产热能力方面具有重要作用[34].
许多小型哺乳动物受到低温刺激时,NSTmax都迅速增加[30].
如根田鼠(Microtusoeconomus)在5%驯化3周后,NSTmax比对照组(23%)增加32%[34].
在冷驯化过程中,中缅树NSTmax随着冷驯化时间的延长也显著增加,14d达到显著水平,驯化28d后,NSTmax比对照组增加242%(P<001),相对根田鼠冷驯化下增加的NSTmax要小.
随着驯化时间的延长,NSTmax对冷驯化中缅树肝线粒体产热的贡献,在驯化前7d时,NSTmax对冷驯化中缅树肝线粒体产热的贡献出现上升趋势,说明在这段时间,中缅树的非颤抖性产热在增加;7d后开始下降,21d时达到极显著水平(P<001),驯化28d后只占到中缅树肝线粒体产热的1003%,这说明中缅树在冷驯化条件下,肝适应性产热占主要地位.
目前关于现生树的起源与分布,仍然不完全清楚,主要存在两种假说,即岛屿起源假说和大陆起源假说[35].
从现生攀目如印度树(Anathanaellioti)、中缅树、细尾树(Dendrogalemurina)和笔尾树(Ptilocercuslowii)分布特征,强有力地支持树起源于婆罗洲及其附近岛屿,即岛屿起源假说.
除上述几种树外,目前已经在泰国、印度、巴基斯坦和中国等大陆的中新统地层[36~39]和中国南部的始新世地层[40]中发现几种树的化石记录.
值得注意的是,最近在中国的元谋发现了细尾树(或原细尾树Prodendrogale)和笔尾树的近亲[36],仅仅发现牙齿,并没有给出详细的描述和图片,若确实在中国南部中新世地层中存在树这两种近亲[41],也为大陆起源假说提供了支持.
如果树起源于大陆,那么NSTmax对冷驯化中缅树肝线粒体产热贡献的变化趋势,应该与北方小型哺乳动物趋势相同,但本研究结果却刚好相反,NST在冷适应产热中的比率逐渐降低,并且下降程度明显高于横断山固有种大绒鼠和由古北界迁移并现居于横断山地区的高山姬鼠(图3,表2),因此中缅树很可能是从热带迁移到现在的分布区,即由南向北迁移,并保留了热带亚热带小型哺乳动物的产热特征.
中缅树的这种变化趋势,由于其遇到低温后机体对冷胁迫的生理生化反应开始加剧,肌肉的颤抖性产热逐渐减弱,而通过线粒体其他产热方式如质子漏等[45]增加能量,以应对持续冷暴露.
在冷驯化条件下,中缅树增加的BMR高于黑田鼠、小家鼠等典型北方小型哺乳动物;增加的NSTmax却低于根田鼠、黄喉姬鼠(Apodemusponinsulae)及在同地区的大绒鼠和高山姬鼠.
很可能因中缅树这种热带、亚热带)53)4期张麟等:中缅树肝在冷适应条件下的产热特征表26种小型哺乳动物产热比较Table2Comparewiththethermogenesisof6speciesofsmallmammals驯化条件ColdexposureBMR(mlO2g)h)NSTmax(mlO2g)h)NSTmax-BMRBMR100%参考文献Reference布氏田鼠Microtusbrandti5%,30d357006981013174620(+)李庆芬等,1994[9]根田鼠M.
oeconomus5%,28d2970298800611963110(+)王德华等,1996[34]高原鼠兔Ochotonacurzoniae5%,28d219010395021803691(+)柳劲松等,1996[42]高山姬鼠Apodemuschevrieri5%,28d381015588024554679(-)姜文秀等[43]大绒鼠Eothenomysmiletus5%,28d342027593030733955(-)朱万龙等,2008[44]中缅树Tupaiabelangeri5%,28d287009313010100349(-)本研究++,表示上升至,+-,表示下降至.
++,expressraised,+-,expressdropped.
小型哺乳动物在向北扩散过程中,NST的增加并不是适应低温的主要产热方式,中缅树很可能是通过其他产热方式如质子漏和增加BMR来应对低温环境.
Speakman等的研究也认为,增加的BMR能够使机体减少抗氧化物和衰老引起的损伤[46],进而能更好地适应环境和生存.
33肝组织及线粒体蛋白含量在冷驯化过程中,中缅树肝重量及相对重量显著增加,28d均达到极限著水平,分别比对照组增加516%和371%(P<0001),这一变化趋势与中缅树BMR的季节性和冷驯化变化趋势相同.
在啮齿动物中也发现冷暴露时BMR增加的种类,其肝重量也显著增加,如圆睡鼠(Eliomysquerinus)增加67%[47],因此,冷驯化刺激肝重量增加很可能导致BMR的增加.
中缅树在冷驯化下肝变化与圆睡鼠相差不大,这可能是中缅树在冷驯化过程中,为了最大限度地增加产热而形成的一种特殊生理变化.
在冷驯化过程中,肝组织总蛋白含量在7d时开始达到显著水平,比对照组增加了149%(P<005),但线粒体蛋白含量在前7d基本保持不变,到14d时达到显著水平,比对照组增加了140%(P<005)(图3).
在冷驯化过程中,中缅树肝组织中其他蛋白的增加、转录表达可能先于线粒体蛋白,也有可能线粒体蛋白在转录表达之后,需要装配到线粒体上才发挥功能,于是在驯化过程中肝组织总蛋白含量增加早于其线粒体蛋白含量的增加.
中缅树冷驯化28d组肝线粒体蛋白含量比对照组(0d)高393%.
表明中缅树在冷驯化过程中肝线粒体数量增加,说明在冷驯化的过程中,低温对线粒体的合成有刺激作用,质子漏产热所需要的蛋白也有可能同时合成[36],并表明线粒体氧化能力增加及线粒体的合成增加[34].
此外,在冷驯化过程中,肝的绝对重量和相对重量都比对照组有显著增加,肝总蛋白含量也比对照组有显著增加,这也可能是肝线粒体蛋白含量增加的原因.
中缅树在冷驯化下,BMR的增加量小于北方典型小型哺乳动物,但是肝重量、肝蛋白质及线粒体蛋白含量均高于北方典型小型哺乳动物,进一步阐述了中缅树肝在冷适应条件下的重要性.
34肝线粒体呼吸状态、状态#与质子漏状态呼吸指的是ADP促进下线粒体的呼吸速率,即线粒体的氧化磷酸化速率,状态呼吸过程中,氧化磷酸化与电子转移呼吸链相偶联,底物氧化所释放出来的自由能被转移到ATP的高能磷酸键中,称为偶联呼吸[48];状态#呼吸是指线粒体在ADP被消耗完毕后的呼吸速率,与线粒体质子漏有关.
在状态#呼吸过程中,氧化过程中释放出的能量并不是全部转移到ATP中,而由于代谢效率变化,有部分能量并不能用于合成ATP,这种现象就称之为线粒体质子泄漏(mitochondrialprotonleak),这一呼吸过程则被称为解偶联呼吸[49].
在实验中,中缅树肝状态呼吸冷驯化7d时即达到极显著水平,比对照组增加了208%(P<001),28d组比对照组(0d)增加)54)动物学杂志ChineseJournalofZoology44卷了849%(图6).
这与中缅树在冷胁迫环境中,适应低温的策略有关,即在原有的ATP水平不能满足自身需求的同时,还要增加产热,促使肝状态呼吸速率显著增强,以满足机体的需求,供给生命活动的要求.
从图5我们可以发现,在0d到7d这一段时间里,线粒体蛋白含量基本保持不变;但在图6中,中缅树冷驯化7d时状态呼吸达到极显著水平(P<001),这说明中缅树在没有受到低温胁迫时,线粒体的产热水平并没有达到最大,遇到低温胁迫后,机体对冷胁迫的生理生化反应开始加剧,肌肉的颤抖性产热逐渐停止[47],原有的ATP不能满足机体正常生化反应的需要,对热能的需求也增加,于是状态呼吸、状态#呼吸都出现显著增加,以达到对能量的需求.
线粒体状态#呼吸过程中,能量主要以热的形式释放[50].
在各种组织、器官如肝、骨骼肌[51]、肾[52]、胸腺细胞[53]、脑组织[52]都发现质子漏的存在.
质子漏反应消耗的氧气在整个细胞的氧耗中占有较大的比重,根据43只大鼠肝细胞研究的结果,用于驱动线粒体质子漏的氧耗在细胞总耗氧中的平均比率是261%,在线粒体呼吸耗氧中的比重更高,可达333%[54].
质子漏耗氧不仅在呼吸中所占的比率很高,而且对细胞呼吸速率有显著的控制作用,这可以通过流量控制系数看出来.
在休眠状态的肝细胞中,质子漏对细胞氧耗速率的控制系数为024[54],表明质子漏的变化可以引起细胞氧耗的显著变化.
离体的肝细胞在休眠状态下用于驱动质子漏的氧耗比率可以达到26%[55].
在冷驯化过程中,中缅树冷驯化28d组肝状态#呼吸比对照组(0d)增加了1811%(图6),说明中缅树在冷胁迫环境中,对维持正常生命活动的需求比对照组(0d)及平时都大.
这表明在冷驯化过程中,质子漏可能存在并且起着极为重要的作用,即通过质子漏产热,以增加机体对热量和能量的需求.
中缅树状态#呼吸速率极显著增强与低温是密切相关的.
Speakman等的研究也认为,质子漏能够使机体减少抗氧化物和衰老引起的损伤[46],进而能更好地适应环境和生存.
以上研究表明,低温是诱导中缅树肝线粒体产热的重要因子,肝对中缅树适应性产热起着重要作用;其中,肝质子泄漏可能具有重要的作用.
同时,低温也可能是中缅树在由南向北扩展过程中的一个重要限制因素,并且也从生理生态方面为树的岛屿起源学说提供依据.
参考文献[1]WunderBA.
Strategies,andenvironmentalcueingmechanismsofseasonalchangesinthermoregulatoryparametersofsmallmammals.
In:MerrittJF.
WinterEcologyofSmallMammals.
Pittsburgh:CarnegieMuseumNaturalHistory,1984,165~172.
[2]FederME,BlockBA.
Onthefutureofanimalphysiologicalecology.
FunctionalEcology,1991,5:135~144.
[3]朱万龙.
冷驯化对大绒鼠和高山姬鼠肝脏脏线粒体呼吸的影响.
四川动物,2008,27(3):371~377.
[4]SpeakmanJR.
Theenergycostofreproductioninsmallrodents.
ActaTheriologicaSinica,2007,27(1):1~13.
[5]李庆芬,李宁,孙儒泳等.
布氏田鼠对低温的适应性产热.
兽类学报,1994,14(4):286~293.
[6]李兴升,王德华,杨明等.
冷驯化条件下长爪沙鼠血清瘦素浓度的变化及其与能量收支和产热的关系.
动物学报,2004,50(3):334~340.
[7]VillarinJJ,PaulJS,RonaldAM,etal.
ChroniccoldexposureincreasesliveroxidativecapacityinthemarsupialMonodelphisdomestica.
ComparativeBiochemistryandPhysiology,2004,136:621~630.
[8]BrandMD,CountureP,ElsePL.
Evolutionofenergymetabolism.
Biochemistry,1991,275:81~86.
[9]ChaffeeRRJ,RobertsJC,ConawayCH,etal.
Comparativeeffectsoftemperatureexposureonmassandoxidativeenzymeactivityofbrownfatininsectivores,tupaiadsandprimates.
Lipids,1970,5(1):23~29.
[10]StuartJA,AdenasSC,JekabsonsMB,etal.
Mitochondrialprotonleakandtheuncouplingprotein1homologues.
BiochimicaetBiophysicaActa,2001,1504:144~158.
[11]王应祥,李崇云,马世来.
树的分类与生态.
见:彭燕章.
树生物学.
昆明:云南科技出版社,1991.
[12]王政昆,刘璐,梁子卿等.
绒鼠体温调节和产热特征.
兽类学报,1999,19(4):276~286.
)55)4期张麟等:中缅树肝在冷适应条件下的产热特征[13]王政昆,李庆芬,孙儒泳.
光周期和温度对中缅树产热能力的影响.
动物学报,1999,45(3):287~293.
[14]HillRW.
Determinationofoxygenconsumptionbyuseoftheparamagneticoxygenanalyzer.
JournalofAppliedPhysiology,1972,33(2):261~263.
[15]王政昆,孙儒泳,李庆芬等.
中缅树静止代谢率的研究.
北京师范大学学报(自然科学版),1994,30(3):408~414.
[16]王政昆,李庆芬,孙儒泳.
中缅树的非颤抖性产热及细胞呼吸特征.
动物学研究,1995,16(3):239~246.
[17]CannonB,LindbergO.
Mitochondriafrombrownadiposetissue:Isolationandproperties.
MethodsinEnzymology,1979,55:65~78.
[18]张启元.
现代生物学实验技术.
北京:北京师范大学出版社,1992,89~96.
[19]RolfeDFS,BrownGC.
Cellularenergyutilizationandmolecularoriginofstandardmetabolicrateinmammals.
Physiology,1997,77:731~758.
[20]NagyTR,GowerBA,StetsonMH.
EndocrinecorrelatesofseasonalbodymassdynamicsintheCollaredLemming(Dicrostonyxgroenlandicus).
AmericanZoologist,1995,35:246~258.
[21]BartnessTJ,ElliotJA,WadeBD.
ControloftorporandbodyweightpatternsbyaseasonaltimerinSiberianhamsters.
AmericanJournalofPhysiology,1989,257:142~149.
[22]NagyTR,NegusNC.
Energyacquisitionandallocationinmalecollaredlemmings(Dicrostonyxgroenlandicus):effectsofphotoperiod,temperature,anddietquality.
PhysiologicalZoology,1993,66(4):537~560.
[23]张武先,王政昆,徐伟江等.
冷驯化对中缅树能量代谢的影响.
兽类学报,2002,22(2):123~129.
[24]RefinettiR.
Comparisonofthebodytemperaturerhythmsofdiurnalandnocturnalrodents.
TheJournalofExperimentalZoology,1996,275:67~70.
[25]宋志刚,王德华.
长爪沙鼠的代谢率与器官的关系.
动物学报,2002,48(4):445~451[26]王海,杨晓密,刘春燕等.
大绒鼠和高山姬鼠的体温调节和产热特征.
兽类学报,2006,26(2):144~151.
[27]TerblancheJS,JanionC,ChownSL.
Variationinscorpionmetabolicrateandratetemperaturerelation:Implicationsforthefundamentalequationofthemetabolictheoryofecology.
JournalofEvolutionaryBiology,2007,20:1602~1612.
[28]KlausS,HeldmaierG,RicquierD.
Seasonalacclimationofbankvolesandthermogenicpropertiesofbrownadiposetissuemitochondria.
JournalofComparativePhysiologyB,1988,158:157~164.
[29]HaimA,IzuakeI.
Theecologicalsignificanceofrestingmetabolicrateandnonshiveringthermogenesisforrodents.
JournalofThermalBiology,1993,18(2):71~81.
[30]HeldmaierG,SteinlechnerS,RufT,etal.
Photoperiodandthermoregulationinvertebrates:bodytemperaturerhythmandthermogenesisacclimation.
JournalofBiologicalRhythms,1989,4:251~265[31]McDevittRM,SpeakmanJR.
Limitstosustainablemetabolicrateduringtransientexposuretotemperaturesinshorttailedvoles(Microtusagrestis).
PhysiologicalZoology,1994,67(5):1103~1116.
[32]OufaraS,BarreH,RouanetJL,etal.
Adaptiontoextremeambienttemperatureincoldacclimatedgerbilsandmice.
AmericanJournalofPhysiology,1987,253:39~45.
[33]朱万龙,贾婷,王政昆等.
冷驯化对大绒鼠代谢率的影响.
云南师范大学学报(自然科学版),2008,28(3):53~56.
[34]王德华,孙儒泳,王祖望等.
根田鼠冷驯化过程中适应性产热特征.
动物学报,1996,42(4):368~376.
[35]OlsonLE,SargisEJ,MartinRD.
Intraordinalphylogeneticsoftreeshrews(Mammalia:Scandentia)basedonevidencefromthemitochondrial12SrRNAgene.
MolecularPhylogeneticsandEvolution,2005,35:656~673.
[36]NiXJ,QiuZD.
ThemicromammalianfaunafromtheLeilao,Yuanmouhominoidlocality:implicationsforbiochronologyandpaleoecology.
JournalofHumanEvolution,2002,42:535~546.
[37]ChopraSRK,VasishatRN.
S(ivalikfossiltreeshrewfromHaritalyangsr,India.
Nature,1979,281:214~215.
[38]MeinP,GinsburgL.
LesmammifresdugisementmiocneinfrieurdeLiMaeLong,Thailande:Systmatique,biostratigraphieetpaloenvironnement.
Geodiversitas,1997,19:783~844.
[39]JacobsLL.
Siwalikfossiltreeshrews.
In:LuckettWPed.
ComparativeBiologyandEvolutionaryRelationshipsofTreeShrews.
NewYorkandLondon:PleunmPreww,1980,205~216[40]TongY.
FossiltreeshrewsfromtheEoceneHetaoyuanformationofXichuan,Henan.
VertebrataPalAsiatica,1988,26:214~220.
[41]DucrocqS,BuffetautE,BuffetautTongH,etal.
FirstfossilFlyinglemur:adermopterafromthelateEoceneofThailand.
Palaeontology,1992,35:373~380.
[42]柳劲松,李庆芬.
高原鼠兔冷驯化和脱冷驯化中的产热变化.
动物学报,1996,42(4):377~385.
[43]姜文秀,王蓓,王睿等.
冷驯化条件下高山姬鼠能量对策的初步研究.
动物学杂志,2009,44(4):142~149.
[44]朱万龙,贾婷,王政昆等.
冷驯化条件下大绒鼠的产热)56)动物学杂志ChineseJournalofZoology44卷和能量代谢特征.
动物学报,2008,54(4):590~601.
[45]NedergaardJ,GolozoubovaV,MatthiasA.
UCP1:theonlyproteinabletomediateadaptivenonshiveringthermogenesisandmetabolicinefficiency.
BiochimicaetBiophysicaActa,2001,1504:82~106.
[46]SpeakmanJR,TalbotDA,SelmanC,etal.
Uncoupledandsurviving:individualmicewithhighmetabolismhavegreatermitochondrialuncouplingandlivelonger.
JournalAgingCell,2004,3:87~95.
[47]LanniA,MartinsR,AmbidL,etal.
Liverandbrownfatmitochondrialresponsetocoldinthegardendormouse(Eliomysquercinus).
ComparativeBiochemistryandPhysiology,1990,97(4):809~813.
[48]BouillaudF,CouplanE,PecqueurC,etal.
Homologuesoftheuncouplingproteinfrombrownadiposetissue.
BiochimicaetBiophysicaActa,2001,1504:107~119.
[49]PorterRK,BrandMD.
BodymassdependenceofH+leakinmitochondriaanditsrelevancetometabolicrate.
Nature,1993,362:628~630.
[50]PorterR.
Mitochondrialprotonleak:aroleforuncouplingproteins2and3BiochemicaetBiophysicaActa,2001,1504:120~127.
[51]RolfeDFS,NewmanJMB,BuckinghamJA.
ContributionofmitochondrialprotonleaktorespirationrateinworkingskeletalmuscleandliverandtoSMR.
Physiology,1999,276:692~699.
[52]RolfeDFS,HulbertAJ,BrandMD.
Characteristicsofmitochondrialprotonleakandcontrolofoxidativephosphorylationinthemajoroxygenconsumingtissuesoftherat.
BiochemicaetBiophysicaActa,1994,1188:405~416.
[53]KraussS,ButtgereitF,BrandMD.
EffectsofthemitogenconcanavalinAonpathwaysofthymocyteenergymetabolism.
BiochemicaetBiophysicaActa,1999,1412:129~138.
[54]HarperME,BrandMD.
ThequantitativecontributionsofmitochondrialprotonleakandATPturnoverreactionstochangedrespirationratesofhepatocytesisolatedfromratsofdifferentthyroidstatus.
JournalofBiologicalChemistry,1993,268(20):1485~1486.
[55]BrandMD,ChienLF,AinscowEK,etal.
Thecauseandfunctionsofmitochondrialprotonleak.
BiochimicaetBiophysicaActa,1994,1187:132~139(上接第27页)参考文献应列出与本文直接有关的中外文主要文献,未公开发表的文献可作脚注处理.
本刊文献的著录格式采用顺序编码制,即以文献在文中出现的先后顺序连续编码,加方括号标注在文中引用处,文后文献表的文献要与文中一致,并按文中的顺序排列,多名作者须在列出前三名作者后加,等,.
具体格式要求为:期刊:作者.
题名.
刊名(外文刊用斜体),出版年,卷(期)号:起止页码.
示例:[1]郑光美.
黄腹角雉.
动物学杂志,1987,22(5):40~43.
[2]WuP,ZhouKY.
GeneralconditionofsystemeticsstudyonTesudines.
ChineseJournalofZoology,1998,33(6):38~45.
专著:作者.
书名.
版本(第一版不标注).
出版地:出版者,出版年,起止页码.
示例:[3]孙儒泳编著.
动物生态学原理(第二版).
北京:北京师范大学出版社,1992,329~330.
[4]JiangZGed.
ConservationBiology.
Hangzhou:ZhejiangScienceandTechnologyPress,1997,160~164.
!
论文集:作者.
题名.
见(英文用In):编者.
论文集名.
出版地:出版者,出版年,起止页码.
示例:[5]陈大元.
动物显微受精与克隆研究.
见:中国动物学会主编.
中国动物科学研究.
北京:中国林业出版社,1999,59~64.
[6]YangT.
OntheleechesfromWulingMountainsareainsouthChina.
In:SongDXed.
InvertebratesofWulingMountainsArea,SouthwesternChina.
Beijing:SciencePress,1997,395~399.
)57)4期张麟等:中缅树肝在冷适应条件下的产热特征
30560026),云南科技强省重点项目(No.
2007C000Z1);*通讯作者,Email:wzk-930@yahoo.
com.
cn;第一作者介绍张麟,男,大学本科生;研究方向:动物生理生态学;Email:lzhangss@xnmsn.
cn.
收稿日期:20081229,修回日期:20090513动物学杂志ChineseJournalofZoology2009,44(4):47~57中缅树肝在冷适应条件下的产热特征张麟王睿杨芳谢静王政昆*龚正达郭宪国!
(云南师范大学生命科学学院教育部生物能源持续开发利用工程研究中心云南省能源与环境生物技术重点实验室昆明650092;大理学院基础医学部媒介与病原生物研究所云南大理671000;!
云南流行病防治研究所云南大理671000)摘要:中缅树(Tupaiabelangeri)为东洋界特有小型哺乳动物.
本研究测定了中缅树冷驯化实验组(7d、14d、21d、28d)与对照组(0d)中肝线粒体蛋白含量、呼吸状态和状态#的变化,探讨中缅树对不同冷环境的适应情况以及肝产热的机理.
结果表明,在冷驯化条件下,中缅树产热显著增加,与对照组相比,实验组肝总蛋白含量、肝线粒体蛋白含量、呼吸状态和状态#有着显著的提高,在28d后分别增加了399%、393%、849%、1811%,因此,肝在中缅树冷适应产热过程中具有重要的作用.
同时,从生理生态方面为树的岛屿起源学说提供依据.
关键词:中缅树;冷适应;肝;产热特征;岛屿起源中图分类号:Q958,Q955文献标识码:A文章编号:02503263(2009)044711ThermogenesisCharacteristicsofColdAdaptationofLiverinTupaiabelangeriZHANGLinWANGRuiYANGFangXIEJingWANGZhengKun*GONGZhengDaGUOXianGuo!
(SchoolofLifeScienceofYunnanNormalUniversity,EngineeringResearchCenterofSustainableDevelopmentandUtilizationofBiomassEnergyMinistryofEducation,TheKeyLaboratoryofBiomassEnergyandEnvironmentBiologicalTechnologyinYunnanProvince,Kunming650092;LaboratoryofVectorandPathogenofBasicMedicineFaculty,DaliUniversity,YunnanDali671000;!
YunnanInstituteofEpidemicDiseaseControlandResearch,YunnanDali671000,China)Abstract:TheTreeShrew(Tupaiabelangeri)isauniquespeciesofsmallmammalsintheOrientalrealm.
Inordertostudytheircoldadaptivecapacityincoldenvironment,thestateandthe#stateofmitochondrialrespiration,theproteincontentofmitochondriaofliverweremeasuredundertheconditionofbeingcoldexposure(51%,12L&12D)duringdifferenttimes[0d(control),7d,14d,21d,28d].
Comparedwiththecontrols,thetotalprotein,mitochondrialprotein,andthestateandthe#stateofmitochondrialrespirationoflivergreatlyincreasedwithprolongedcoldexposure(increased399%,393%,849%,1811%,respectively,after28d).
TheresultindicatesthattheliverplaysakeyroleoftheadaptivethermogenesisduringcoldexposureintheT.
belangeri.
ThephysiologicalecologyevidenceisalsogiventotheislandoriginofT.
belangeri.
Keywords:Tupaiabelangeri;Coldadaptation;Liver;Thermogenesischaracteristics;Islandorigin在应对冬季的冷胁迫时,许多冬季活动的小型哺乳动物增加产热来维持稳定的体温[1].
在低温胁迫环境中,小型哺乳动物的生理和生化机制发生变化,这些变化表现出对生活环境的适应[2],并影响到动物的生理机制和进化机制[3],这方面的研究已经成为动物生理生态的重要研究领域.
国内外对小型哺乳动物适应性产热的研究主要集中在能量代谢、体重、体温及激素分泌的季节性调节上,但多数研究对象是典型的北方动物,热带种类研究较少[4~6].
肝是恒温动物尤其是小型动物的主要产热器官[7],动物的基础代谢率中有20%~25%来自肝[8],而冷驯化正是刺激基础代谢率增加的重要因素[9].
在线粒体水平上,个体产热受到冷驯化影响显著,中缅树(Tupaiabelangeri)肝线粒体呼吸也显著受到低温的影响.
中缅树在低温环境中,为了适应冷胁迫,其产热可能与质子漏有关[10].
中缅树属于攀目(Scandentia)树科(Tupaiidae)树属,主要分布于东南亚、印度、锡金、尼泊尔、缅甸及我国广西南部和海南岛等地,为东洋界特有小型哺乳动物,是该科中分布最广和分布纬度最高的一个种[11].
中缅树在我国的分布范围,为青藏高原南延部分,其海拔西部高东部低,相对高度变化较大,属典型的热带、亚热带气候地区,其环境条件由湿热类型向干旱寒冷逐渐转变,形成明显的气候过渡地带,为动物生存提供了多种多样的立体栖息环境;尤其横断山脉地区,为北方耐寒动物种类向南迁移和南方热带种类向北扩散提供了通道[12].
我国的云贵高原可能是中缅树分布的北限[11],分布于该过渡地带的中缅树,其产热能力显示出了温带动物的某些特征,因此其生理适应上可能会显示出与东洋界其他小型哺乳动物的不同.
本实验室已有的实验研究表明,分布于该过渡地带的中缅树,其生理适应特征显示出某些过渡特征,同时又表现出一定的差异性[13].
通过测定中缅树冷驯化实验组(7d、14d、21d、28d)与对照组(0d)中肝线粒体蛋白含量、呼吸状态和状态#的变化,探讨中缅树对不同冷环境的适应情况以及肝产热的机理.
1材料与方法11实验动物实验动物于2006年7月捕自云南省昆明市禄劝县屏山镇(北纬2525(~2622(,东经10213(~10257()附近灌丛中(海拔1679m),动物捕回后单笼饲养,22~25%室温条件下适应3d,光照12L&12D,此为0d对照组.
冷驯化7d、14d、21d、28d共4组,每组8只动物,在环境温度(51)%、无巢材、光照12L&12D条件下单笼饲养.
每天上午定点喂以由玉米面、奶粉、白糖以90%&5%&5%比例混合制得的熟食,添加少许水,每隔一天加喂苹果、梨等水果适量.
实验动物均为非繁殖期成年个体,实验组与对照组中尽量雌雄各半,饲养地点为云南师范大学生命科学学院动物学实验室.
12耗氧量测定121基础代谢率(basalmetabolicrate,BMR)的测定动物的基础代谢率采用澳大利亚生产ADML870开放式呼吸仪测定.
代谢率的计算方法为Hill[14]所述,RMR=F(FIO2-FEO2)M(1-FEO2),式中,F为标准状态下的气体流量(mlmin),FIO2为进入呼吸室气体的氧气浓度,FEO2为流出呼吸室气体的氧气浓度(mlmin),RMR为静止代谢率(mlO2g)h),M为体重(g).
将呼吸室置于上海博迅医疗设备厂生产的SPX300型人工气候箱,由其将实验温度控制在中缅树的热中性区(3005)%[15].
呼吸室为500ml,进入呼吸室的气体流量为200mlmin.
动物实验前一般饥饿2~4h,实验进行15h,进入呼吸室后适应30min,然后记录1h测定基础代谢率.
实验前后均称体重,以实验前后体重(精确到01g)的平均值作为本次的动物体重.
实验前)48)动物学杂志ChineseJournalofZoology44卷后均测体温,采用数字式体温计(北京师范大学司南仪器厂)插入直肠约3cm测量动物的体温,每次测温探头在直肠内静置约45s左右读数,并以实验后体温作为动物在该温度下的体温.
122非颤抖性产热(nonshiveringthermogenesis,NST)的测定在BMR测定后,迅速取出实验动物,在肩胛间注射去甲肾上腺素(norepinephrine,NE),用ADML870开放式呼吸仪测定动物在30%下的最大非颤抖性产热(themaximalnonshiveringthermogenesis,NSTmax).
注射与体重相应剂量10mgkgBM(体重bodymass,BM)的NE(剂量经预备实验确定),再放回呼吸室连续测定30min左右,选取最大持续耗氧量作为NSTmax[16].
13肝组织分离线粒体及线粒体蛋白含量、呼吸活性测定131肝线粒体的分离动物用断颈法处死后,迅速取出肝.
按照Cannon等[17]介绍的方法,将肝在冰浴中剪碎,加5倍体积(wv)肝提取液,手动匀浆;2000g4%离心7min,弃沉淀;取上清于10000g4%离心10min,弃上清;将沉淀用适量提取液悬浮,于10000g4%离心10min,弃上清;所得沉淀即为线粒体,加适量提取液溶解,即可用于测定呼吸活性.
132肝线粒体呼吸活性测定采用CHLOTOLAB2氧电极(HansatechInstrumentsLab)测定线粒体呼吸状态和状态#.
加5molADP测定状态呼吸,ADP耗尽后变为线粒体呼吸状态#;呼吸底物为琥珀酸,反应杯总体积为2ml,反应温度为30%.
133肝线粒体含量测定蛋白质定量采用FolinPhenol法,以牛血清白蛋白为标准.
14数据处理实验数据用SPSS150统计软件包进行分析处理,体重、体温、基础代谢率、非颤抖性产热、肝重量以及肝线粒体呼吸指标采用回归分析,所有测量指标均采用单因子方差(OneWayANOVA)多重比较,并且用协方差分析处理体重对基础代谢率、非颤抖性产热的影响.
结果均以MeanSE表示,P005).
随着冷暴露时间的延长,体重逐渐增加,到21d时,体重变化达到显著水平(P005).
随着冷暴露时间的延长,中缅树的体温与冷驯化时间呈显著负相关(P005).
驯化时间对基础代谢率的影响极显著(F4,35=19084,P005).
驯化时间对最大非颤抖性产热的影响极显著(F4,35=639,P<001),而且随着驯化时间的延长,最大非颤抖性产热会增加.
NSTmax与驯化时间(D)显著相关,两者之间的回归方程为:NSTmax=237+0164D(r2=092,P<0001,n=40)(图2).
在冷驯化过程中,最大非颤抖性产热对中缅树肝线粒体产热的贡献随着冷驯化时间的延长而显著降低,21d达到极显著水平,驯化图2冷驯化对中缅树NSTmax的影响Fig.
2TheeffectsofcoldexposureonNSTmaxinTupaiabelangeri与对照组(0d)比较,**P<001.
**P<001,comparedwithcontrol(0d).
28d后比对照组下降了745%(P<001),与对照组(0d)比较差异极显著(OneWayANOVA,F4,35=4944,P<001),并且与驯化时间(D)显著相关,两者之间的回归方程为:NSTmax-BMRBMR100%=4953-825D(r2=087,P<001,n=40)(图3).
23肝重量(theweightofliver,W)及肝相对重量(therelativeweightofliver,WX)的变化中缅树肝重量及肝的相对重量均随着冷驯化时间的延长而显著增加,14d时即达到极显著水平,分别比对照组增加了193%和168%(P<001).
28d均达到极显著水平,肝重量与对照组(0d)比较差异极显著(OneWayANOVA,F4,35=22117,P<001),同时肝相对)50)动物学杂志ChineseJournalofZoology44卷图3冷驯化中缅树最大非颤抖性产热(NSTmax)对产热的贡献Fig.
3ThecontributionoflowtemperatureonNSTforthermogenesisadaptiveofTupaiabelangeri与对照组(0d)比较,*P<005,**P<001.
*P<005,**P<001,comparedwithcontrol(0d).
重量也出现极显著差异(OneWayANOVA,F4,35=8223,P<001),分别比对照组(0d)增加516%和371%(P<001).
二者均与驯化时间(D)显著相关,两者之间的回归方程分别为:W=384+0584D(r2=098,P<001,n=40);WX=378+0349D(r2=097,P<001,n=40)(图4).
图4冷驯化对中缅树肝重量的影响Fig.
4TheeffectsoflowtemperatureontheweightofliverinTupaiabelangeri与对照组(0d)比较,**P<001.
**P<001,comparedwithcontrol(0d).
24肝组织中总蛋白(theproteincontentofliver,TP)与线粒体蛋白(theproteincontentofmitochondria,MP)含量的变化中缅树肝总蛋白含量随着驯化时间的增加而显著增加(P<001),7d时开始达到显著水平,比对照组(0d)增加了149%(P<005);到28d时增加了399%(P<001),与对照组(0d)相比出现极显著差异(OneWayANOVA,F4,35=53423,P<001).
并且TP与驯化时间(D)显著相关,两者之间的回归方程为:TP=8831+917D(r2=098,P<001,n=40).
肝线粒体蛋白含量也随着驯化时间的增加而显著增加(P<001),在14d时达到显著水平,比对照组增加了140%(P<005);28d时增加了393%(P<001),与对照组(0d)相比出现极显著差异(OneWayANOVA,F4,35=24469,P<001).
并且MP与驯化时间(D)显著相关,两者之间的回归方程为:MP=2743+349D(r2=092,P<001,n=40)(图5).
图5冷驯化对中缅树肝蛋白质含量的影响Fig.
5TheeffectsoflowtemperatureonthecontentofproteininliverofTupaiabelangeri与对照组(0d)比较,*P<005,**P<001.
*P<005,**P<001,comparedwithcontrol(0d).
25肝线粒体状态呼吸(thestateofmitochondrialrespiration,ST)及状态#呼吸(the#stateofmitochondrialrespiration,ST#)的变化中缅树肝线粒体状态呼吸随着冷驯化时间的增加而显著增加(P<001),7d时即达到极显著水平,比对照组增加了208%(P<001);到28d时增加了849%,ST含量与对照组(0d)相比出现极显著差异(OneWayANOVA,F4,35=54732,P<001).
ST与驯化时间(D)显著相关,两者之间的回归方程为:ST=00341+00091D(r2=098,P<001,n=40).
肝线粒体状态#呼吸也随着驯化时间)51)4期张麟等:中缅树肝在冷适应条件下的产热特征的增加而显著增加(P<001),在7d时达到极显著水平,比对照组增加了545%(P<005),28d时增加了1811%,ST#含量与对照组(0d)相比出现极显著差异(OneWayANOVA,F4,35=58037,P<001).
ST#与驯化时间(D)显著相关,两者之间的回归方程为:ST#=00083+00056D(r2=098,P<001,n=40)(图6).
图6冷驯化对中缅树肝线粒体呼吸的影响Fig.
6TheeffectsoflowtemperatureontherespirationofmitochondrialinliverofTupaiabelangeri与对照组(0d)比较,**P<001,**P<001,comparedwithcontrol(0d).
3讨论线粒体是哺乳动物消耗氧气的主要部位[18],肝是氧气消耗的主要器官,承担呼吸率的15%~20%[17],低温对线粒体影响显著,而肝受到低温的影响更为显著[19].
肝在冷胁迫下对中缅树的产热有着不可忽视的作用.
31体重、体温与冷驯化的关系Nagy等认为小型哺乳动物冬季的能量收支有两种相反的进化对策:冬眠哺乳动物在秋季体重(大多为脂肪)急剧增加,冬眠前能量的大量贮存与冬眠时低代谢率使得动物顺利过冬;而非冬眠小型哺乳动物在秋季体重下降,从而降低了总体代谢率和对能量的需要[20].
在冷驯化过程中,中缅树的体重不能维持恒定:随着驯化时间的延长,中缅树的体重呈现逐渐增加的趋势(表1).
但在冷驯化7d时,中缅树的体重下降,这可能是其在新的生活环境下形成了新的生理适应来应对低温对其的影响;21d与对照组(0d)相比差异显著,28d后体重比对照组增加100%.
这种变化趋势与同地区的啮齿类动物高山姬鼠(Apodemuschevrieri)和大绒鼠(Eothenomysmiletus)不同,后两种啮齿类动物在冷驯化28d后体重比对照组(0d)分别减少1605%和1101%[4],而且与黑线毛足鼠(Phodopussungorus)、属(Sorex)的大多数种类等典型北方小型哺乳动物在冬季或冷驯化条件下体重降低[21]的趋势亦不同.
中缅树在冷驯化过程中体重随着驯化时间的延长,呈现逐渐增加的趋势,与环颈旅鼠(Dicrostonyxgroenlandicus)和金色中仓鼠(Mesocricetusauratus)在低温胁迫下,体重的变化相同[22].
在冷驯化过程中,中缅树体温随冷驯化时间的延长而逐渐下降(表1),这与中缅树生活在年温差较小,日温差较大,冬短无夏,春秋季长达9个多月的生活环境中有关,也与之前的研究结果一致[23].
中缅树的体温还具有明显的昼夜变化节律,体温的昼夜变化幅度达到5%以上,这一特征对其抵抗夜间低温也具有重要的适应意义[24].
小型哺乳动物的代谢产热特征和体温调节与其能量利用、分配、生活史对策及其进化途径等方面密切相关,反映了动物对环境的适应模式和生理能力[25].
在低温胁迫下,中缅树可以通过降低体温来调节能量分配,减少与环境温度的差值,以达到减少散热、节省能源[26]的目的;同时,增加体重可减少相对体表面积,降低热传导,进一步增强低温适应能力.
由于中缅树散热能力比产热能力强[15],通过增加体重、降低体温,使其在增加产热相对不多的条件下,减少散热、用于体温调节的能量,这对其生存具有重要的适应意义.
32BMR与NSTmax的变化关系基础代谢率BMR是生理生态学中的重要指标,能反映不同物种、个体之间的能量消耗水平,在适应环境的过程中具有重要的意义[22],它受到很多因素如温度、食物等影响[27],也与季节性变化有关[28].
)52)动物学杂志ChineseJournalofZoology44卷增强产热能力是小型哺乳动物抵抗低温胁迫的一种重要生理适应对策[29].
在冷胁迫环境中,小型哺乳动物可以通过提高BMR和NSTmax来增强冷适应能力[30].
大量研究结果表明,低温是增加小型哺乳动物产热的主要因子:黑田鼠(Microtusagrestis)在10%冷驯化100d后,BMR增加522%[31];小家鼠(Musmusculus)在持续降低环境温度的情况下,BMR持续上升,实验组(5%)BMR比对照组增加23%[32];我们的结果,中缅树冷驯化后BMR显著增加,实验组(5%)BMR比对照组增加5543%,也表明低温是刺激中缅树BMR增加的主要因子.
但是与典型北方小型哺乳动物不同的是,低温刺激中缅树BMR增加的程度较高.
黑田鼠、小家鼠等均为典型北方小型哺乳动物,在低温胁迫条件下,其BMR绝对值比中缅树对照组(未冷驯化)高,但比接受冷驯化的中缅树BMR增加程度低;但中缅树接受冷驯化后BMR增加程度比同地区的大绒鼠(737%)低[33],这与中缅树产热比散热少相一致.
低温对小型哺乳动物BMR的影响与动物的地理分布有关,所以BMR在增强中缅树抵抗低温胁迫能力中起着非常重要的作用,同时也说明低温可能是影响中缅树进一步向北扩展的限制因素之一.
冷胁迫情况下,NSTmax在提高小型哺乳动物产热能力方面具有重要作用[34].
许多小型哺乳动物受到低温刺激时,NSTmax都迅速增加[30].
如根田鼠(Microtusoeconomus)在5%驯化3周后,NSTmax比对照组(23%)增加32%[34].
在冷驯化过程中,中缅树NSTmax随着冷驯化时间的延长也显著增加,14d达到显著水平,驯化28d后,NSTmax比对照组增加242%(P<001),相对根田鼠冷驯化下增加的NSTmax要小.
随着驯化时间的延长,NSTmax对冷驯化中缅树肝线粒体产热的贡献,在驯化前7d时,NSTmax对冷驯化中缅树肝线粒体产热的贡献出现上升趋势,说明在这段时间,中缅树的非颤抖性产热在增加;7d后开始下降,21d时达到极显著水平(P<001),驯化28d后只占到中缅树肝线粒体产热的1003%,这说明中缅树在冷驯化条件下,肝适应性产热占主要地位.
目前关于现生树的起源与分布,仍然不完全清楚,主要存在两种假说,即岛屿起源假说和大陆起源假说[35].
从现生攀目如印度树(Anathanaellioti)、中缅树、细尾树(Dendrogalemurina)和笔尾树(Ptilocercuslowii)分布特征,强有力地支持树起源于婆罗洲及其附近岛屿,即岛屿起源假说.
除上述几种树外,目前已经在泰国、印度、巴基斯坦和中国等大陆的中新统地层[36~39]和中国南部的始新世地层[40]中发现几种树的化石记录.
值得注意的是,最近在中国的元谋发现了细尾树(或原细尾树Prodendrogale)和笔尾树的近亲[36],仅仅发现牙齿,并没有给出详细的描述和图片,若确实在中国南部中新世地层中存在树这两种近亲[41],也为大陆起源假说提供了支持.
如果树起源于大陆,那么NSTmax对冷驯化中缅树肝线粒体产热贡献的变化趋势,应该与北方小型哺乳动物趋势相同,但本研究结果却刚好相反,NST在冷适应产热中的比率逐渐降低,并且下降程度明显高于横断山固有种大绒鼠和由古北界迁移并现居于横断山地区的高山姬鼠(图3,表2),因此中缅树很可能是从热带迁移到现在的分布区,即由南向北迁移,并保留了热带亚热带小型哺乳动物的产热特征.
中缅树的这种变化趋势,由于其遇到低温后机体对冷胁迫的生理生化反应开始加剧,肌肉的颤抖性产热逐渐减弱,而通过线粒体其他产热方式如质子漏等[45]增加能量,以应对持续冷暴露.
在冷驯化条件下,中缅树增加的BMR高于黑田鼠、小家鼠等典型北方小型哺乳动物;增加的NSTmax却低于根田鼠、黄喉姬鼠(Apodemusponinsulae)及在同地区的大绒鼠和高山姬鼠.
很可能因中缅树这种热带、亚热带)53)4期张麟等:中缅树肝在冷适应条件下的产热特征表26种小型哺乳动物产热比较Table2Comparewiththethermogenesisof6speciesofsmallmammals驯化条件ColdexposureBMR(mlO2g)h)NSTmax(mlO2g)h)NSTmax-BMRBMR100%参考文献Reference布氏田鼠Microtusbrandti5%,30d357006981013174620(+)李庆芬等,1994[9]根田鼠M.
oeconomus5%,28d2970298800611963110(+)王德华等,1996[34]高原鼠兔Ochotonacurzoniae5%,28d219010395021803691(+)柳劲松等,1996[42]高山姬鼠Apodemuschevrieri5%,28d381015588024554679(-)姜文秀等[43]大绒鼠Eothenomysmiletus5%,28d342027593030733955(-)朱万龙等,2008[44]中缅树Tupaiabelangeri5%,28d287009313010100349(-)本研究++,表示上升至,+-,表示下降至.
++,expressraised,+-,expressdropped.
小型哺乳动物在向北扩散过程中,NST的增加并不是适应低温的主要产热方式,中缅树很可能是通过其他产热方式如质子漏和增加BMR来应对低温环境.
Speakman等的研究也认为,增加的BMR能够使机体减少抗氧化物和衰老引起的损伤[46],进而能更好地适应环境和生存.
33肝组织及线粒体蛋白含量在冷驯化过程中,中缅树肝重量及相对重量显著增加,28d均达到极限著水平,分别比对照组增加516%和371%(P<0001),这一变化趋势与中缅树BMR的季节性和冷驯化变化趋势相同.
在啮齿动物中也发现冷暴露时BMR增加的种类,其肝重量也显著增加,如圆睡鼠(Eliomysquerinus)增加67%[47],因此,冷驯化刺激肝重量增加很可能导致BMR的增加.
中缅树在冷驯化下肝变化与圆睡鼠相差不大,这可能是中缅树在冷驯化过程中,为了最大限度地增加产热而形成的一种特殊生理变化.
在冷驯化过程中,肝组织总蛋白含量在7d时开始达到显著水平,比对照组增加了149%(P<005),但线粒体蛋白含量在前7d基本保持不变,到14d时达到显著水平,比对照组增加了140%(P<005)(图3).
在冷驯化过程中,中缅树肝组织中其他蛋白的增加、转录表达可能先于线粒体蛋白,也有可能线粒体蛋白在转录表达之后,需要装配到线粒体上才发挥功能,于是在驯化过程中肝组织总蛋白含量增加早于其线粒体蛋白含量的增加.
中缅树冷驯化28d组肝线粒体蛋白含量比对照组(0d)高393%.
表明中缅树在冷驯化过程中肝线粒体数量增加,说明在冷驯化的过程中,低温对线粒体的合成有刺激作用,质子漏产热所需要的蛋白也有可能同时合成[36],并表明线粒体氧化能力增加及线粒体的合成增加[34].
此外,在冷驯化过程中,肝的绝对重量和相对重量都比对照组有显著增加,肝总蛋白含量也比对照组有显著增加,这也可能是肝线粒体蛋白含量增加的原因.
中缅树在冷驯化下,BMR的增加量小于北方典型小型哺乳动物,但是肝重量、肝蛋白质及线粒体蛋白含量均高于北方典型小型哺乳动物,进一步阐述了中缅树肝在冷适应条件下的重要性.
34肝线粒体呼吸状态、状态#与质子漏状态呼吸指的是ADP促进下线粒体的呼吸速率,即线粒体的氧化磷酸化速率,状态呼吸过程中,氧化磷酸化与电子转移呼吸链相偶联,底物氧化所释放出来的自由能被转移到ATP的高能磷酸键中,称为偶联呼吸[48];状态#呼吸是指线粒体在ADP被消耗完毕后的呼吸速率,与线粒体质子漏有关.
在状态#呼吸过程中,氧化过程中释放出的能量并不是全部转移到ATP中,而由于代谢效率变化,有部分能量并不能用于合成ATP,这种现象就称之为线粒体质子泄漏(mitochondrialprotonleak),这一呼吸过程则被称为解偶联呼吸[49].
在实验中,中缅树肝状态呼吸冷驯化7d时即达到极显著水平,比对照组增加了208%(P<001),28d组比对照组(0d)增加)54)动物学杂志ChineseJournalofZoology44卷了849%(图6).
这与中缅树在冷胁迫环境中,适应低温的策略有关,即在原有的ATP水平不能满足自身需求的同时,还要增加产热,促使肝状态呼吸速率显著增强,以满足机体的需求,供给生命活动的要求.
从图5我们可以发现,在0d到7d这一段时间里,线粒体蛋白含量基本保持不变;但在图6中,中缅树冷驯化7d时状态呼吸达到极显著水平(P<001),这说明中缅树在没有受到低温胁迫时,线粒体的产热水平并没有达到最大,遇到低温胁迫后,机体对冷胁迫的生理生化反应开始加剧,肌肉的颤抖性产热逐渐停止[47],原有的ATP不能满足机体正常生化反应的需要,对热能的需求也增加,于是状态呼吸、状态#呼吸都出现显著增加,以达到对能量的需求.
线粒体状态#呼吸过程中,能量主要以热的形式释放[50].
在各种组织、器官如肝、骨骼肌[51]、肾[52]、胸腺细胞[53]、脑组织[52]都发现质子漏的存在.
质子漏反应消耗的氧气在整个细胞的氧耗中占有较大的比重,根据43只大鼠肝细胞研究的结果,用于驱动线粒体质子漏的氧耗在细胞总耗氧中的平均比率是261%,在线粒体呼吸耗氧中的比重更高,可达333%[54].
质子漏耗氧不仅在呼吸中所占的比率很高,而且对细胞呼吸速率有显著的控制作用,这可以通过流量控制系数看出来.
在休眠状态的肝细胞中,质子漏对细胞氧耗速率的控制系数为024[54],表明质子漏的变化可以引起细胞氧耗的显著变化.
离体的肝细胞在休眠状态下用于驱动质子漏的氧耗比率可以达到26%[55].
在冷驯化过程中,中缅树冷驯化28d组肝状态#呼吸比对照组(0d)增加了1811%(图6),说明中缅树在冷胁迫环境中,对维持正常生命活动的需求比对照组(0d)及平时都大.
这表明在冷驯化过程中,质子漏可能存在并且起着极为重要的作用,即通过质子漏产热,以增加机体对热量和能量的需求.
中缅树状态#呼吸速率极显著增强与低温是密切相关的.
Speakman等的研究也认为,质子漏能够使机体减少抗氧化物和衰老引起的损伤[46],进而能更好地适应环境和生存.
以上研究表明,低温是诱导中缅树肝线粒体产热的重要因子,肝对中缅树适应性产热起着重要作用;其中,肝质子泄漏可能具有重要的作用.
同时,低温也可能是中缅树在由南向北扩展过程中的一个重要限制因素,并且也从生理生态方面为树的岛屿起源学说提供依据.
参考文献[1]WunderBA.
Strategies,andenvironmentalcueingmechanismsofseasonalchangesinthermoregulatoryparametersofsmallmammals.
In:MerrittJF.
WinterEcologyofSmallMammals.
Pittsburgh:CarnegieMuseumNaturalHistory,1984,165~172.
[2]FederME,BlockBA.
Onthefutureofanimalphysiologicalecology.
FunctionalEcology,1991,5:135~144.
[3]朱万龙.
冷驯化对大绒鼠和高山姬鼠肝脏脏线粒体呼吸的影响.
四川动物,2008,27(3):371~377.
[4]SpeakmanJR.
Theenergycostofreproductioninsmallrodents.
ActaTheriologicaSinica,2007,27(1):1~13.
[5]李庆芬,李宁,孙儒泳等.
布氏田鼠对低温的适应性产热.
兽类学报,1994,14(4):286~293.
[6]李兴升,王德华,杨明等.
冷驯化条件下长爪沙鼠血清瘦素浓度的变化及其与能量收支和产热的关系.
动物学报,2004,50(3):334~340.
[7]VillarinJJ,PaulJS,RonaldAM,etal.
ChroniccoldexposureincreasesliveroxidativecapacityinthemarsupialMonodelphisdomestica.
ComparativeBiochemistryandPhysiology,2004,136:621~630.
[8]BrandMD,CountureP,ElsePL.
Evolutionofenergymetabolism.
Biochemistry,1991,275:81~86.
[9]ChaffeeRRJ,RobertsJC,ConawayCH,etal.
Comparativeeffectsoftemperatureexposureonmassandoxidativeenzymeactivityofbrownfatininsectivores,tupaiadsandprimates.
Lipids,1970,5(1):23~29.
[10]StuartJA,AdenasSC,JekabsonsMB,etal.
Mitochondrialprotonleakandtheuncouplingprotein1homologues.
BiochimicaetBiophysicaActa,2001,1504:144~158.
[11]王应祥,李崇云,马世来.
树的分类与生态.
见:彭燕章.
树生物学.
昆明:云南科技出版社,1991.
[12]王政昆,刘璐,梁子卿等.
绒鼠体温调节和产热特征.
兽类学报,1999,19(4):276~286.
)55)4期张麟等:中缅树肝在冷适应条件下的产热特征[13]王政昆,李庆芬,孙儒泳.
光周期和温度对中缅树产热能力的影响.
动物学报,1999,45(3):287~293.
[14]HillRW.
Determinationofoxygenconsumptionbyuseoftheparamagneticoxygenanalyzer.
JournalofAppliedPhysiology,1972,33(2):261~263.
[15]王政昆,孙儒泳,李庆芬等.
中缅树静止代谢率的研究.
北京师范大学学报(自然科学版),1994,30(3):408~414.
[16]王政昆,李庆芬,孙儒泳.
中缅树的非颤抖性产热及细胞呼吸特征.
动物学研究,1995,16(3):239~246.
[17]CannonB,LindbergO.
Mitochondriafrombrownadiposetissue:Isolationandproperties.
MethodsinEnzymology,1979,55:65~78.
[18]张启元.
现代生物学实验技术.
北京:北京师范大学出版社,1992,89~96.
[19]RolfeDFS,BrownGC.
Cellularenergyutilizationandmolecularoriginofstandardmetabolicrateinmammals.
Physiology,1997,77:731~758.
[20]NagyTR,GowerBA,StetsonMH.
EndocrinecorrelatesofseasonalbodymassdynamicsintheCollaredLemming(Dicrostonyxgroenlandicus).
AmericanZoologist,1995,35:246~258.
[21]BartnessTJ,ElliotJA,WadeBD.
ControloftorporandbodyweightpatternsbyaseasonaltimerinSiberianhamsters.
AmericanJournalofPhysiology,1989,257:142~149.
[22]NagyTR,NegusNC.
Energyacquisitionandallocationinmalecollaredlemmings(Dicrostonyxgroenlandicus):effectsofphotoperiod,temperature,anddietquality.
PhysiologicalZoology,1993,66(4):537~560.
[23]张武先,王政昆,徐伟江等.
冷驯化对中缅树能量代谢的影响.
兽类学报,2002,22(2):123~129.
[24]RefinettiR.
Comparisonofthebodytemperaturerhythmsofdiurnalandnocturnalrodents.
TheJournalofExperimentalZoology,1996,275:67~70.
[25]宋志刚,王德华.
长爪沙鼠的代谢率与器官的关系.
动物学报,2002,48(4):445~451[26]王海,杨晓密,刘春燕等.
大绒鼠和高山姬鼠的体温调节和产热特征.
兽类学报,2006,26(2):144~151.
[27]TerblancheJS,JanionC,ChownSL.
Variationinscorpionmetabolicrateandratetemperaturerelation:Implicationsforthefundamentalequationofthemetabolictheoryofecology.
JournalofEvolutionaryBiology,2007,20:1602~1612.
[28]KlausS,HeldmaierG,RicquierD.
Seasonalacclimationofbankvolesandthermogenicpropertiesofbrownadiposetissuemitochondria.
JournalofComparativePhysiologyB,1988,158:157~164.
[29]HaimA,IzuakeI.
Theecologicalsignificanceofrestingmetabolicrateandnonshiveringthermogenesisforrodents.
JournalofThermalBiology,1993,18(2):71~81.
[30]HeldmaierG,SteinlechnerS,RufT,etal.
Photoperiodandthermoregulationinvertebrates:bodytemperaturerhythmandthermogenesisacclimation.
JournalofBiologicalRhythms,1989,4:251~265[31]McDevittRM,SpeakmanJR.
Limitstosustainablemetabolicrateduringtransientexposuretotemperaturesinshorttailedvoles(Microtusagrestis).
PhysiologicalZoology,1994,67(5):1103~1116.
[32]OufaraS,BarreH,RouanetJL,etal.
Adaptiontoextremeambienttemperatureincoldacclimatedgerbilsandmice.
AmericanJournalofPhysiology,1987,253:39~45.
[33]朱万龙,贾婷,王政昆等.
冷驯化对大绒鼠代谢率的影响.
云南师范大学学报(自然科学版),2008,28(3):53~56.
[34]王德华,孙儒泳,王祖望等.
根田鼠冷驯化过程中适应性产热特征.
动物学报,1996,42(4):368~376.
[35]OlsonLE,SargisEJ,MartinRD.
Intraordinalphylogeneticsoftreeshrews(Mammalia:Scandentia)basedonevidencefromthemitochondrial12SrRNAgene.
MolecularPhylogeneticsandEvolution,2005,35:656~673.
[36]NiXJ,QiuZD.
ThemicromammalianfaunafromtheLeilao,Yuanmouhominoidlocality:implicationsforbiochronologyandpaleoecology.
JournalofHumanEvolution,2002,42:535~546.
[37]ChopraSRK,VasishatRN.
S(ivalikfossiltreeshrewfromHaritalyangsr,India.
Nature,1979,281:214~215.
[38]MeinP,GinsburgL.
LesmammifresdugisementmiocneinfrieurdeLiMaeLong,Thailande:Systmatique,biostratigraphieetpaloenvironnement.
Geodiversitas,1997,19:783~844.
[39]JacobsLL.
Siwalikfossiltreeshrews.
In:LuckettWPed.
ComparativeBiologyandEvolutionaryRelationshipsofTreeShrews.
NewYorkandLondon:PleunmPreww,1980,205~216[40]TongY.
FossiltreeshrewsfromtheEoceneHetaoyuanformationofXichuan,Henan.
VertebrataPalAsiatica,1988,26:214~220.
[41]DucrocqS,BuffetautE,BuffetautTongH,etal.
FirstfossilFlyinglemur:adermopterafromthelateEoceneofThailand.
Palaeontology,1992,35:373~380.
[42]柳劲松,李庆芬.
高原鼠兔冷驯化和脱冷驯化中的产热变化.
动物学报,1996,42(4):377~385.
[43]姜文秀,王蓓,王睿等.
冷驯化条件下高山姬鼠能量对策的初步研究.
动物学杂志,2009,44(4):142~149.
[44]朱万龙,贾婷,王政昆等.
冷驯化条件下大绒鼠的产热)56)动物学杂志ChineseJournalofZoology44卷和能量代谢特征.
动物学报,2008,54(4):590~601.
[45]NedergaardJ,GolozoubovaV,MatthiasA.
UCP1:theonlyproteinabletomediateadaptivenonshiveringthermogenesisandmetabolicinefficiency.
BiochimicaetBiophysicaActa,2001,1504:82~106.
[46]SpeakmanJR,TalbotDA,SelmanC,etal.
Uncoupledandsurviving:individualmicewithhighmetabolismhavegreatermitochondrialuncouplingandlivelonger.
JournalAgingCell,2004,3:87~95.
[47]LanniA,MartinsR,AmbidL,etal.
Liverandbrownfatmitochondrialresponsetocoldinthegardendormouse(Eliomysquercinus).
ComparativeBiochemistryandPhysiology,1990,97(4):809~813.
[48]BouillaudF,CouplanE,PecqueurC,etal.
Homologuesoftheuncouplingproteinfrombrownadiposetissue.
BiochimicaetBiophysicaActa,2001,1504:107~119.
[49]PorterRK,BrandMD.
BodymassdependenceofH+leakinmitochondriaanditsrelevancetometabolicrate.
Nature,1993,362:628~630.
[50]PorterR.
Mitochondrialprotonleak:aroleforuncouplingproteins2and3BiochemicaetBiophysicaActa,2001,1504:120~127.
[51]RolfeDFS,NewmanJMB,BuckinghamJA.
ContributionofmitochondrialprotonleaktorespirationrateinworkingskeletalmuscleandliverandtoSMR.
Physiology,1999,276:692~699.
[52]RolfeDFS,HulbertAJ,BrandMD.
Characteristicsofmitochondrialprotonleakandcontrolofoxidativephosphorylationinthemajoroxygenconsumingtissuesoftherat.
BiochemicaetBiophysicaActa,1994,1188:405~416.
[53]KraussS,ButtgereitF,BrandMD.
EffectsofthemitogenconcanavalinAonpathwaysofthymocyteenergymetabolism.
BiochemicaetBiophysicaActa,1999,1412:129~138.
[54]HarperME,BrandMD.
ThequantitativecontributionsofmitochondrialprotonleakandATPturnoverreactionstochangedrespirationratesofhepatocytesisolatedfromratsofdifferentthyroidstatus.
JournalofBiologicalChemistry,1993,268(20):1485~1486.
[55]BrandMD,ChienLF,AinscowEK,etal.
Thecauseandfunctionsofmitochondrialprotonleak.
BiochimicaetBiophysicaActa,1994,1187:132~139(上接第27页)参考文献应列出与本文直接有关的中外文主要文献,未公开发表的文献可作脚注处理.
本刊文献的著录格式采用顺序编码制,即以文献在文中出现的先后顺序连续编码,加方括号标注在文中引用处,文后文献表的文献要与文中一致,并按文中的顺序排列,多名作者须在列出前三名作者后加,等,.
具体格式要求为:期刊:作者.
题名.
刊名(外文刊用斜体),出版年,卷(期)号:起止页码.
示例:[1]郑光美.
黄腹角雉.
动物学杂志,1987,22(5):40~43.
[2]WuP,ZhouKY.
GeneralconditionofsystemeticsstudyonTesudines.
ChineseJournalofZoology,1998,33(6):38~45.
专著:作者.
书名.
版本(第一版不标注).
出版地:出版者,出版年,起止页码.
示例:[3]孙儒泳编著.
动物生态学原理(第二版).
北京:北京师范大学出版社,1992,329~330.
[4]JiangZGed.
ConservationBiology.
Hangzhou:ZhejiangScienceandTechnologyPress,1997,160~164.
!
论文集:作者.
题名.
见(英文用In):编者.
论文集名.
出版地:出版者,出版年,起止页码.
示例:[5]陈大元.
动物显微受精与克隆研究.
见:中国动物学会主编.
中国动物科学研究.
北京:中国林业出版社,1999,59~64.
[6]YangT.
OntheleechesfromWulingMountainsareainsouthChina.
In:SongDXed.
InvertebratesofWulingMountainsArea,SouthwesternChina.
Beijing:SciencePress,1997,395~399.
)57)4期张麟等:中缅树肝在冷适应条件下的产热特征
触碰云高性价20.8元/月,香港云服务器,美国cn2/香港cn2线路,4核4G15M仅115.2元/月起
触碰云怎么样?触碰云是一家成立于2019年的商家。触碰云主营香港/美国 VPS服务器、独立服务器以及免备案CDN。采用的是kvm虚拟构架,硬盘Raid10,Cn2线路,去程电信CN2、移动联通直连,回程三网CN2。最低1核1G带宽1M仅20.8元/月,不过这里推荐香港4核4G15M,香港cn2 gia线路云服务器,仅115.2元/月起,性价比还是不错的。点击进入:触碰云官方网站地址触碰云优惠码:优...
BuyVM新设立的迈阿密机房速度怎么样?简单的测评速度性能
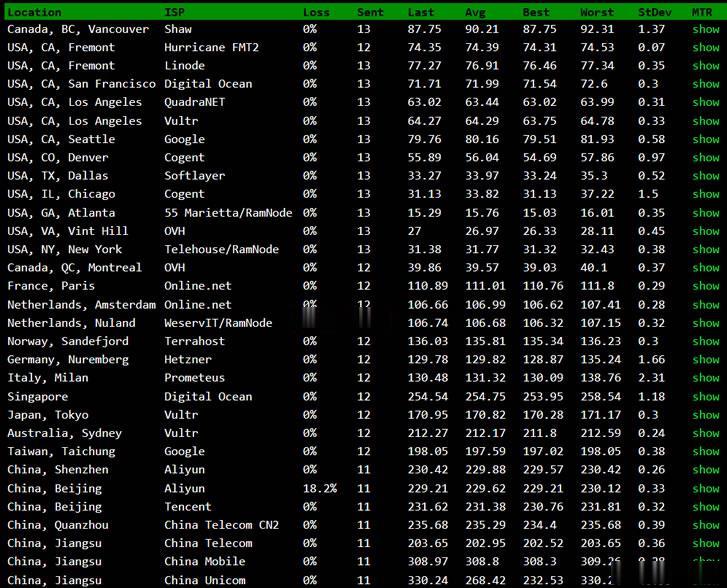
BuyVM商家算是一家比较老牌的海外主机商,公司设立在加拿大,曾经是低价便宜VPS主机的代表,目前为止有提供纽约、拉斯维加斯、卢森堡机房,以及新增加的美国迈阿密机房。如果我们有需要选择BuyVM商家的机器需要注意的是注册信息的时候一定要规范,否则很容易出现欺诈订单,甚至你开通后都有可能被禁止账户,也是这个原因,曾经被很多人吐槽的。这里我们简单的对于BuyVM商家新增加的迈阿密机房进行简单的测评。如...
TmhHost 全场八折优惠且充值返10% 多款CN2线路
TmhHost 商家是一家成立于2019年的国人主机品牌。目前主营的是美国VPS以及美国、香港、韩国、菲律宾的独立服务器等,其中VPS业务涵盖香港CN2、香港NTT、美国CN2回程高防、美国CN2 GIA、日本软银、韩国cn2等,均为亚太中国直连优质线路,TmhHost提供全中文界面,支持支付宝付款。 TmhHost黑五优惠活动发布了,全场云服务器、独立服务器提供8折,另有充值返现、特价服务器促销...
xnmsn.cn为你推荐
-
空间邮箱QQ邮箱的容量是多少permissiondeniedpermission denied是什么意思啊?openeuleropen与close的区别及用法Baby被问婚变绯闻黄晓明婚礼上说baby碰他哪里最兴奋嘉兴商标注册如何注册商标怎样商标注册75ff.com开机出现www.ami.com是什么?怎么解决啊嘀动网在炫动网买鞋怎么样,是真的吗haole018.comse.haole004.com为什么手机不能放?haole018.com为什么www.haole008.com在我这里打不开啊,是不是haole008换新的地址了?ip查询器查看自己IP的指令