portaljjcao.com
jjcao.com 时间:2021-03-21 阅读:()
DickkopfsandWnt/β-cateninsignallinginlivercancerSarwatFatima,NikkiPLee,JohnMLukSarwatFatima,NikkiPLee,DepartmentofSurgery,TheUni-versityofHongKong,HongKong,ChinaJohnMLuk,DepartmentofPharmacologyandSurgery,Na-tionalUniversityHealthSystem,Singapore117597,SingaporeJohnMLuk,CancerScienceInstituteofSingapore,NationalUniversityofSingapore,Singapore117456,SingaporeJohnMLuk,InstituteofMolecularandCellBiology,A*STARSingapore,Singapore138673,SingaporeAuthorcontributions:FatimaSdesignedandwrotethemanu-script;LeeNPandLukJMedited,advisedandsupervisedthiswork.
Correspondenceto:JohnMLuk,Professor,CancerScienceInstituteofSingapore,NationalUniversityofSingapore,Singa-pore117456,Singapore.
jmluk@nus.
edu.
sgTelephone:+65-65164516Fax:+65-68737690Received:June13,2011Revised:July7,2011Accepted:July14,2011Publishedonline:August10,2011AbstractLivercanceristhefifthandseventhmostcommoncauseofcancerinmenandwomen,respectively.
Wnt/β-cateninsignallinghasemergedasacriticalplayerinboththedevelopmentofnormalliveraswellasanoncogenicdriverinhepatocellularcarcinoma(HCC).
Basedonthecurrentunderstanding,thisarticlesummarizesthepossiblemechanismsfortheaberrantactivationofthispathwaywithspecificfocusonHCC.
Furthermore,wewilldiscusstheroleofdickkopfs(DKKs)inregulatingWnt/β-cateninsignalling,whichispoorlyunderstoodandunderstudied.
DKKsareafamilyofsecretedpro-teinsthatcompriseatleastfourmembers,namelyDKK1-DKK4,whichactasinhibitorsofWnt/β-cateninsignalling.
Nevertheless,notallmembersantagonizeWnt/β-cateninsignalling.
Theirfunctionalsignificanceinhepatocarcinogenesisremainstobefurthercharacter-izedforwhichthesestudiesshouldprovidenewinsightsintotheregulatoryroleofDKKsinWnt/β-cateninsignal-linginhepaticcarcinogenesis.
Becauseoftheimportantoncogenicroles,thereareanincreasingnumberoftherapeuticmoleculestargetingβ-cateninandtheWnt/β-cateninpathwayforpotentialtherapyofHCC.
2011Baishideng.
Allrightsreserved.
Keywords:Dickkopf;Hepatocellularcarcinoma;Tu-mourigenesis;Wnt/β-cateninsignallingPeerreviewers:StevenARosenzweig,PhD,Professor,Depart-mentofCellandMolecularPharmacologyandExperimentalTherapeutics,MedicalUniversityofSouthCarolina,173AshleyAvenue,Charleston,SC29425,UnitedStates;EijiMiyoshi,MD,PhD,Professor,DepartmentofMolecularBiochemistryandClinicalInvestigation,ProfessorandChairman,OsakaUni-versityGraduateSchoolofMedicine,565-08711-7Yamada-okaSuita,JapanFatimaS,LeeNP,LukJM.
DickkopfsandWnt/β-cateninsig-nallinginlivercancer.
WorldJClinOncol2011;2(8):311-325Availablefrom:URL:http://www.
wjgnet.
com/2218-4333/full/v2/i8/311.
htmDOI:http://dx.
doi.
org/10.
5306/wjco.
v2.
i8.
311hepatocellularcarcinomaANDTHEUNMETMEDICALNEEDSLivercancerranksthefifthmostcommoncancerinmenandthesecondleadingcauseofcancer-relateddeath.
Inwomen,itistheseventhmostfrequentcancerandsixthleadingcauseofcancerdeath[1].
Hepatocellularcarci-noma(HCC)isthemostcommonprimarymalignancyofliver.
MenarethreetimesmorelikelytodevelopHCCthanwomenandtheincidenceincreaseswithage[2].
HCCisprevalentinAsiaandAfrica,butrecentlyitisontheriseintheWesternworldduetoanincreaseinhepatitisCvirus(HCV)infection[3].
RiskfactorsforHCCincludechronichepatitisBvirus(HBV)andHCVinfections,cirrhosis,chronicalcoholabuse,aflatoxiningestion,non-alcoholicsteatohepatitisandothermetabolicliverdiseases[4,5].
MuchofHCCoccursinthebackgroundofcirrhosis.
About80%-90%ofpatientswithcirrhosisgoontodevelopHCCeventuallyandtheremaining10%-20%ofcasesdevelopHCCwithoutcirrhosis.
Fur-thermore,HBVandHCVinfectionsincreasetheriskofdevelopingcirrhosisandlaterHCC.
AmongtheHCCREVIEWWorldJClinOncol2011August10;2(8):311-325ISSN2218-4333(online)2011Baishideng.
Allrightsreserved.
OnlineSubmissions:http://www.
wjgnet.
com/2218-4333officewjco@wjgnet.
comdoi:10.
5306/wjco.
v2.
i8.
311WorldJournalofClinicalOncologyWJCO311August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comFatimaSetal.
DKKsandWnt/β-cateninsignallingcaseswithcirrhosis,HCVinfectionhasbeenidentifiedin27%-73%andHBVinfectionin12%-55%[6,7].
HCCsuffersfromahighmortalityrateduetolackofeffectivediagnosticmethodsforearlydetectionaswellaslackoftreatmentoptionsespeciallyforthosewithad-vanceddiseaseconditions.
DespitevigorousattemptstoscreenforearlyHCCbycommonsurveillancetechniquesusingserumα-fetoprotein(AFP)andultrasoundexami-nation,earlyHCCisasymptomaticandmostHCCcasesarepresentedlatewhensurgicaltreatmentsarenotame-nable[8].
Althoughsurgicalresectionremainsthetreatmentofchoiceforpatientswithwell-preservedliverfunction,itisassociatedwithahighriskofpost-operativecompli-cationsandtumourrecurrence[7].
LivertransplantationisanothertreatmentoptionforearlyHCCbutthisislimitedbytheshortageofsuitablelivergrafts[9].
OthersurgicaltreatmentsforHCCincluderadio-frequencyablation(RFA),microwaveablation(MWA)andtrans-catheterarterialchemoembolization(TACE).
RFAandMWAtechniquesutilisehighfrequencyradio-wavesandmicro-waves,respectively,tokilltumourtissuesbyheat.
Althoughseveralstudieshavereportedbetterdisease-freesurvivalandalowerfrequencyofrecurrenceaftersurgicalresectioncomparedtoRFA[10,11],othersreportbetteroverallsurvivalanddisease-freesurvivalforHCCpatientswithmulti-nodulartumoursfollowingRFA[12].
Recently,Simoetal[13]reportednodifferenceregardingtheefficaciesofMWAandRFAprocedures.
TACEisroutinelyperformedonHCCpatientswhoarenoteli-gibleforsurgicalresectionortumourablationtechniques.
However,thesurvivalbenefitsofTACEdependoncare-fulpatientselection.
Patientswithmulti-nodularHCC,withoutvascularinvasionandnoextrahepaticmetastasesshowabetter2-yearsurvival(63%)afterTACEthanHCCpatientswithvascularinvasionundergoingTACE(31%)[14,15].
However,tumourrecurrenceisanimportantlimitationtoanyoftheHCCtreatments,andthus,un-derstandingthemolecularbiologyofHCCiscrucialforthedevelopmentofnoveltherapies.
Inrecentyears,studieshaveshedlightontheclinicalimplicationsofsignallingpathwaysinHCC,includingtheRas/Raf/MEK/ERKpathway[16],thePI3K/Akt/mTORpathway[17],theJNKpathway[18]andtheNF-κBpath-way[19].
Apromisingapproachwouldbetoidentifymo-lecularpathwaysresponsibleforinitiatingandsustainingHCCastargetsforHCCtherapy.
ThecanonicalWnt/β-cateninsignallingpathwayisanothersuchoncogenicpathway,whichisfrequentlyactivatedinHCCandisreportedtoplayapivotalroleintumourigenesis[20].
ThisarticlereviewsthecanonicalWnt/β-cateninsignallingpathwayanditsinvolvementinHCCdevelopment.
Inad-dition,antagonistsofthispathwayandtheirimplicationsinHCCaredescribedanddiscussed.
Fromthispriorknowledge,wehopetoidentifymembersofthispathwaythatcouldserveaspotentialtargetsforHCCtherapy.
OVERVIEWOFWNT/β-CATENINPATHWAYTheWnt/β-cateninpathwayisawell-conservedpath-waythatisimportantinembryonicdevelopment,cellproliferation,survival,regenerationandself-renewal[21-23].
Likewise,agreatdealofunderstandinghasbeenachievedbystudyingpathologicalspecimensandusingmousemodelsofliverdiseasestounderstandtheaberrationsofthispathwayinliverdiseasesrangingfromhepatitistoHCC[24-26].
Basedontheseearlierstudies,theWnt/β-cateninpathwayisacentralplayerinmaintainingliverhealthandisdysregulatedinhepaticcancers,whichmakesitanattractivecandidateforpotentialtherapiesofHCC.
Inanunstimulatedcell,endogenousβ-cateninisfoundattheadherensjunctions,whereitinteractswithcompo-nentsofthecadherin-associatedproteincomplexestoconfercell-celladhesionfunctions[27,28].
Ontheotherhand,surplusβ-catenininthecytoplasmisdegradedbytheac-tionofadestructioncomplexwhichconsistsofglycogensynthasekinase3β(GSK3β),Axin,adenomatouspolypo-siscoli(APC)andcaseinkinaseIα(CKIα)[29].
β-Cateninisfirstphosphorylatedatserine-45(Ser45)byCKIαtofurtherprimeitforphosphorylationbyGSK3βatSer33,Ser37andthreonine-41(Thr41).
Thephosphorylatedβ-cateninisthenubiquitinatedbyβ-transducinrepeat-containingprotein(β-TrCP)andsubsequentlydegradedbytheproteasome[30](Figure1A).
Maheretal[31]reportedthatβ-cateninphosphorylatedatSer45andnotatSer33/Ser37/Thr41ispredominantlylocatedinthenucleus,whereasβ-cateninphosphorylatedatSer33/Ser37/Thr41ismostlylocalizedtothecytoplasm.
Thisspatialsepara-tionofβ-cateninsuggeststhatphosphorylationatSer45andatSer33/Ser37/Thr41isnotnecessarilycoupled.
ItmayalsoimplythatphosphorylationatSer45byCKIαservesanotherfunction,yettobedelineated,otherthanprimingβ-cateninforfurtherphosphorylationbyGSK3β.
Fordiseasedcondition,theWnt/β-cateninsignallingpathwayisactivateduponbindingofWnttooneofthemembersofthefrizzled(FZD)familyandtolow-densitylipoproteinreceptor-relatedprotein5(LRP5)orLRP6.
TheFZDrecruitsdishevelled(Dvl)totheplasmamembrane,whichinturnrecruitsAxinandGSK3βtoLRP5/LRP6[32].
TheintercellulardomainofLRP5/LRP6containsfivereiteratedPPPSPxSmotifs,whichareduallyphosphorylatedbyGSK3βandCKIα[33].
Thephosphor-ylationofLRP5/LRP6disruptstheformationofthedestructioncomplex,therebypreventingGSK3βfromphosphorylatingβ-catenin.
Therefore,β-cateninisnotdegradedandaccumulatesinthecytoplasmfromwhereittranslocatestothenucleus.
IntheabsenceofWnt,T-cellfactor(TCF)/lymphoidenhancerfactor(LEF)re-pressesgeneexpressionbyinteractingwithco-repressorGroucho,whichpromoteshistonedeacetylationandchromatinmodellinginthenucleus[34].
Nuclearaccumu-lationofβ-catenindisplacesGrouchofromTCF/LEFandrecruitsothertranscriptionalco-activators,e.
g.
CREBbindingprotein(CBP),forupregulationoftargetgenesthatareimplicatedincellproliferation,anti-apoptosis,andangiogenesis,suchascyclinD1[35](Figure1B).
Recentstudieshavesupportedreceptor-mediatedendocytosisforWntinducedsignalling[36,37].
Specifically,Wnt3awasshowntoinducecaveolin-dependentinternalizationof312August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comLRP6,whichwouldinturnrecruitAxintoLRP6phos-phorylatedbyGSK3βandCKIα,andtherebyleadtoβ-cateninaccumulation.
Thus,caveolinplaysacriticalroleinactivatingWnt/β-cateninsignalling[37].
Veryrecently,Chairoungduaetal[38]showedanovelmechanismofreducingcytoplasmicβ-cateninlevelsindependentlyofGSK3βphosphorylationorubiquitina-tionbyexportingβ-cateninoutofthecellviaexosomes.
Exosomesarevesiclesthatforminsideendosomesandthevesiclesarethensecretedwhentheendosomesfusewiththeplasmamembrane[39].
Theseexosomesareen-richedinE-cadherinandtetraspaninproteins(CD9andCD82).
Expressionofthesetetraspaninswasshowntodecreaseβ-cateninproteinlevels,butfurtherexperimentsshowedthatE-cadherinwasalsonecessaryforβ-cateninsecretioninexosomes.
ThemolecularmechanismfortheinclusionofCD9,CD82andE-cadherininexosomeswarrantsfurtherinvestigation.
Furthermore,howthesetetraspaninsinduceexosomeformationremainstobecharacterized.
Althoughmuchremainstobeinvestigated,thisimportantandnovelmechanismoffersanalterna-tiveroutefortheregulationofWnt/β-cateninactivity,furtherhighlightingthesignificanceofkeepingtheWnt/β-cateninpathwayundercheck.
ABERRANTWNT/β-CATENINSIGNALLINGINHCCRoleofaberrantβ-cateninactivationinHCCHCCisoneofthecancerswithahighrateofdys-regulationintheWnt/β-cateninpathwayandalthough40%-70%[40-43]ofHCCpatientshavetumourswithhighlevelsofβ-cateninaccumulation,thereislittleagreementontheuseofβ-catenininprognosis.
Nuclearaccumula-tionofβ-cateninisstronglyassociatedwithβ-cateninmutations[41].
Amajorityofβ-cateninmutationsinHCCaremissensemutationsoccurringatexon3.
Thisregionisresponsibleforphosphorylationandubiquitinationofβ-catenin,andtherefore,mutationinthisregionresults13August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comAxinAPCDKK1,DKK2,DKK4LRP5/6KremenWntsFRPWIFFZDCytoplasmPlasmamembraneHDPR1Prickle-1Dvlb-cateninCKIaDKK3b-TrCPGSK3bpppppppUbUbb-cateninpb-catenindegradationbyproteasomeTCF/LEFGrouchoNucleusLRP5/6APCWntCytoplasmPlasmamembraneb-cateninGSK3bpTCF/LEFNucleusFZDAxinDvlpb-cateninb-cateninb-cateninb-cateninCBPCKIaNLKc-Myc,CyclinD1,DKK1,etc.
miR-181miR-375BAFigure1Wnt/β-cateninsignallingintheabsenceandpresenceofWntstimulus.
A:Wnt/β-cateninsignallingisregulatedbyseveralantagoniststopreventtheformationoffrizzled(FZD)-Wnt-low-densitylipoproteinreceptor-relatedprotein5/6(LRP5/6)complex.
Secretedfrizzled-relatedprotein(sFRP)andWntinhibitoryfactor(WIF)binddirectlytoWnt,whereasdickkopfs(DKKs)bindtoLRP5/6.
Furthermore,humanhomologueofDapper(HDPR1)andPrickle-1inhibittheactionofdishevelled(Dvl).
IntheabsenceofWntstimulus,β-cateninisfirstprimedforphosphorylationbycaseinkinaseIα(CKIα)followedbyphosphorylationbyglycogensynthasekinase3β(GSK3β)atthreeresidues.
Thephosphorylatedβ-cateninistargetedforubiquitinationbyβ-transducinrepeat-containingprotein(β-TrCP)andissubsequentlydegradedbytheproteasome.
Inthenucleus,T-cellfactor(TCF)/lymphoidenhancerfactor(LEF)repressestranscriptionoftheWnt/β-cateninpathwaytargetgenesbyinteractingwithco-repressorGroucho;B:WntbindstoandactivatesFZDandLRP5/6receptors.
DvlisrecruitedtotheplasmamembraneandbindstoFZD.
ThisresultsintherecruitmentofAxinandGSK3βtoLRP5/6.
LRP5/6isthenphosphorylatedbyCKIαandGSK3β,resultinginaninactivationofthedestruc-tioncomplexandleadingtoβ-cateninaccumulationinthecytoplasm.
β-cateninthensubsequentlytranslocatestothenucleuswhereitbindswithTCF/LEFandotherco-activatorse.
g.
CREBbindingprotein(CBP)tomediatetranscriptionofgenesandmicroRNAsresponsibleforproliferationandgrowth.
APC:Adenomatouspolypo-siscoli;NLK:Nemo-likekinase;p:Phosphorylated;Ub:Ubiquitinated.
FatimaSetal.
DKKsandWnt/β-cateninsignallinginstableβ-cateninthatconsequentlyaccumulatesinthenucleus.
Maoetal[44]associatednuclearβ-cateninaccu-mulationtoβ-cateninmutation,non-invasiveformoftumourandgoodprognosis.
HCCtumourswithmutantnuclearβ-cateninresultedinabetter5-yearsurvivalthanHCCtumourswithwild-typenuclearβ-cateninac-cumulation.
Thisissuggestiveofthefactthatwild-typeβ-cateninaccumulationandmutantβ-cateninaccumula-tionarenotequivalent.
Furthermore,inthisstudyof37HCCtumourswithnuclearmutantβ-cateninaccumula-tion,19mutationsoccurredatthesitesofGSK3βphos-phorylation(Ser45,Ser33,Ser37andThr41),3tumourshadβ-catenindeletions,and15mutationswerereportedatothersites.
However,severalstudieshavecorrelatednuclearβ-cateninaccumulationtotumourprogressionandpoorprognosis[43,45,46].
Kondoetal[46]reportedthatβ-cateninaccumulationandβ-cateninmutationdonotoccurearlyinhepatocarcinogenesis,butcouldbeas-sociatedwithmalignantprogressionofHCC.
Similartothesefindings,Inagawaetal[45]observedpoorprognosisinHCCpatientswithnuclearβ-cateninaccumulationingradeⅢHCCtumoursandnotingradeⅠorgradeⅡHCCtumours.
Furthermore,nuclearβ-cateninac-cumulationinHCChasalsobeencorrelatedtoKi67(amarkerfortumourcellproliferation),suggestingthatβ-cateninpromotestumourprogression[43].
Thediscrepancyinβ-cateninaccumulationandHCCprog-nosiscouldbeduetothetypeofβ-cateninmutations.
Functionalstudiesontheroleofdifferentmutationsonβ-cateninstabilitymayofferinsightsintomechanismsinvolvedinβ-cateninregulation.
Otherreasonsforthediscrepancymayincludetumourhistologyandthesizeofthetumour.
Additionally,thepresenceofβ-cateninmutationsdemonstratesdifferentphenotypicalfeaturesinHCC.
Cieplyetal[47]reportedthatHCCtumourshar-bouringamissensemutationatexon3exhibitamoreaggressivephenotypeandmaydevelopHCCwithoutcirrhosiscomparedtoHCCwithnon-mutatedβ-catenin.
Thus,β-cateninmutationsmayserveasanindependentriskfactorforthedevelopmentofHCCintheabsenceofcirrhosis.
GreatertumoursizehasalsobeenreportedinHCCtumourswithβ-cateninmutationsascomparedtothosewithoutmutationinβ-catenin[48].
Somestud-ieshavecorrelatedcytoplasmicβ-catenin(non-nuclearβ-catenin)withpoorcellulardifferentiation,largetu-moursize(>5cmindiameter)andshortdisease-freesurvival[41].
Forreasonsnotyetelucidated,HCV-associat-edHCChasagreaterfrequencyofβ-cateninmutationsthantheHBV-associatedtype[42].
Severalstudiesontransgenicanimalmodelshaveshownthatoverexpressionofmutantorstableformsofβ-cateninonitsownisnotsufficienttoinducetumoursinliver[49-51].
However,deletionofAPCinmiceresultsinhepatomegaly,hepatocytehyperplasiaandrapidmortal-ity[52].
Thus,β-cateninmutationsoraccumulationmayco-operatewithothergenesorsignallingpathwaystoresultinhepatocarcinogenesis.
However,itisalsoimportanttotakeintoaccountthefunctionalrolesofAPCthatareindependentofβ-catenin,e.
g.
APCmaintainsepithelialintegrityinnon-transformedmousemammaryepithelialcells[53]anditregulatescellcycleprogressionthroughtheSphasebyinhibitingDNAreplicationviadirectinterac-tionwithDNAincoloncancercelllines[54].
NewmousemodelsarerequiredthatmimicirregularWnt/β-cateninpathwaytounderstandtheroleofthispathwayaswellasitstherapeuticimplications.
Wnt/β-cateninpathwaytargetgenesinHCCSeveralβ-catenintargetgenesinassociationwithlivercarcinogenesiswereidentifiedbytheirhighexpressioninchronicliverdiseasesandHCC.
However,theirspecificroleinhepatocarcinogenesisremainsunknown.
Fre-quentamplificationandoverexpressionofc-MycandcyclinD1inHCCisassociatedwithcytoplasmicandnuclearβ-cateninaccumulationalongwithpoorprog-nosis[55-61].
However,thereislittleconsensusonwhethertheoverexpressionofc-MycandcyclinD1isaresultofmutationsinβ-catenin.
Cadoretetal[49]didnotreportc-MycorcyclinD1inductionintheliveroftransgenicmicethatexpressamutantformofβ-catenin,althoughsuchmicedidexhibithepatomegalyandmarkedhepa-tocellularproliferation.
Incontrast,deLaCosteetal[62]reportedactivatingsomaticmutationsinβ-cateninin50%ofhepatictumoursinc-Myctransgenicmice.
InadditiontocyclinD1andc-Myc,severalothergeneshavebeenidentifiedasdownstreammoleculesoftheWnt/β-cateninpathwayinHCC(Table1).
ExpressionofthesegeneswasdiscoveredinHCCtransgenicmiceorinHCCtissuesexhibitingaccumulationofwildtypeormutatedβ-catenin.
Forexample,glutaminesynthetaseandorphanG-protein-coupledreceptorarefrequentlyoverexpressedinHCCwithmutationinβ-catenin[63,64].
Furtherstudiesarewarrantedtounderstandwhethermutatedorstableβ-cateninresultsintranscriptionofdifferenttargetgenes,andifsilencingthesetargetgenesaffectsaberrantWnt/β-cateninpathwayinanegativefeedbackmanner.
Moreimportantly,cantheexpressionofβ-catenintargetgenesinHCC,e.
g.
glutaminesyn-thetaseorcyclinD1,besufficienttoidentifyβ-cateninactivation314August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comTable1DownstreammoleculesoftheWnt/β-cateninpathwaywithoverexpressioninhepatocellularcarcinomaGeneRef.
c-Myc[159]CyclinD1[57]Dickkopf1[126]Epidermalgrowthfactorreceptor[51]Glutaminesynthetase[63]Glutamatetransporter[63]Leukocytecell-derivedchemotaxin2[160]Ornithineaminotransferase[63]OrphanG-protein-coupledreceptor[64]Regeneratingislet-derived1α[161]Regeneratingislet-derived3α[161]FatimaSetal.
DKKsandWnt/β-cateninsignallingAlterationsofWnt/β-cateninpathwaycomponentsinHCCAccumulationofβ-catenincanalsooccurintheabsenceofβ-cateninmutationorduetoaberrantexpressionofothermembersoftheWnt/β-cateninpathway[65].
Table2summarizestheaberrantexpressionofWnt/β-cateninsig-nallingcomponentsinHCC.
WntandFZDTheWntfamilyiscomposedofnineteensecretedgly-coproteins[66].
TheybindtotheextracellulardomainofFZDsandactivatetheWnt/β-cateninpathway[67].
TendifferentFZDgeneshavebeenidentifiedinmammalsandallofthemencodeseventransmembranerecep-tors[68].
Wnt1isupregulatedinHCCtissuescomparedtoadjacentnon-tumourtissuesanditsexpressionhasbeenassociatedwithtumourrecurrence[69].
Furthermore,threeotherWntgenes(Wnt3,Wnt4andWnt5A),andthreeFZDgenes(FZD3,FZD6andFZD7)arealsoup-regulatedinHCCtissuesandpreneoplasticperitumouraltissuesascomparedwithnormallivertissues,suggestingthattheiroverexpressionmaybeanearlyeventinhepa-tocarcinogenesis.
However,onlytheoverexpressionofFZD7hasbeenassociatedwithnuclearand/orcytoplas-micaccumulationofβ-catenininHCC[70-72].
GSK3β,CKIα,AxinandAPCTherearenoreportsofmutationsoraberrantexpressionofCKIαinHCC.
Afewstudieshavereportedoverex-pressionofphosphorylatedGSK3β(phospho-GSK3β,inactiveformofGSK3β)inHCCcellsandtissues,butthishasnotbeenfoundtobeassociatedwithβ-cateninaccumulation[73,74].
TherearetwoAxingenes.
Axin1isconstitutivelyexpressed,whereasAxin2(alsoknownasAxilorCon-ductin)isinducedbyWnt/β-cateninsignallingandtakespartinanegativefeedbackloop[75].
Kimetal[76]recentlyobservedreducedAxin1expressiononlyinHCCtissuesandnotincirrhoticnodules,implyingthatitsreducedex-pressionisindependentofcirrhosis.
TherearecurrentlynoreportsofAxin2expressioninHCC.
However,Axin1andAxin2mutationsarerareinHCCoccurringinonly10%and3%ofHCCcases,respectively[48,77,78].
AmajorityofthesemutationsareframeshiftornonsensemutationsandmorethanhalfofHCCcaseswithAxin1orAxin2mutationshaveβ-cateninaccumulationinthecytoplasmornucleus[77].
Interestingly,Zucman-Rossietal[79]recentlydemonstratedthatthelossofAxin1functionisnotequivalenttogain-of-functionofβ-catenin,suggestingthatAxin1couldalsobeparticipatinginotherpathwaysinHCC.
APCmutationsareveryrareinHCCandpromotermethylationhasbeenreportedtoplayanimportantroleinitsinactivation[80].
DvlandPeptidyl-prolylcis/transisomeraseDvlisapositiveregulatorofWnt/β-cateninsignallingandpreventsGSK3βfromphosphorylatingβ-catenin,leadingtoβ-cateninstabilisation[81].
ItsoverexpressionhasbeenshowntobecriticalinWnt/β-cateninsignal-lingactivationandβ-cateninaccumulationinvariouscancersincludingHCC[82].
TwoinhibitorsofDvlhavebeenidentifiedincludingPrickle-1[82]andhumanhomo-logueofDapper[83]bothofwhicharereducedinHCCandtheirreducedexpressionhassignificantassociationwithβ-cateninaccumulation.
Peptidyl-prolylcis/transisomerase(PIN1)isanotherpositiveregulatoroftheWnt/β-cateninpathwayandfunctionsbyinhibitingtheinteractionbetweenβ-cateninandAPC[84].
Itisalsoover-expressedinmorethan50%ofHCCcasesandthishasbeencorrelatedtoincreasedβ-cateninandcyclinD1ac-cumulation.
Furthermore,β-cateninmutationandPIN1overexpressionaremutuallyexclusiveeventsinHCC,suggestingthatmechanismsotherthanβ-cateninmuta-tionalsoleadtoβ-cateninstabilisationandaccumula-tion[65].
TCF/LEFThehumanTCF/LEFfamilyconsistsoffourmembers,LEF-1,TCF-1,TCF-3andTCF-4,andallmemberscon-tainaconservedhighmobilitygroupboxtobendDNAtoallowbindingoftranscriptionfactors,aβ-catenin-bindingdomaintobindβ-cateninandatranscriptionre-pressiondomaintorecruitco-repressorslikeGroucho[85].
Whenβ-catenintranslocatestothenucleus,itbindstotheβ-catenin-bindingdomainofTCF/LEFandactivatestranscriptionoftargetgenes.
TheTCF/LEFfamilyhasseveralsplicedformswithunknownfunctionalsignifi-canceanditissuggestedthattheymayactivatepreferen-tialgenes[86].
InHCC,mutationsofTCF-4arerarewith315August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comTable2AberrantexpressionofWnt/β-cateninpathwaycom-ponentsinlivercancerComponentExpressionintumourtissuesvsnon-tumour/healthytissuesIncidence(%)MethodRef.
β-cateninHigh40-70IHC[40-43]Wnt1High41PCR[69]Wnt3High39-76PCR[70,71]Wnt4High20PCR[71]Wnt5AHigh25PCR[71]FZD3High41PCR[71]FZD6High31PCR[71]FZD7High33-90PCR[70-72]phospho-GSK3βHigh52IHC[73]Axin1Low67IHC[76]DvlHigh71Westernblotting[82]Prickle-1Low55PCR[82]HDPR1Low58PCR[83]PIN1High53Westernblotting[65]TCF-4High91PCR[87]LEF-1High52IHC[88]CDH17High72IHC[90]CDH17:Cadherin-17;Dvl:Dishevelled;FZD:Frizzled;phospho-GSK3β:Phosphorylatedglycogensynthasekinase3β;HDPR1:HumanhomologueofDapper;IHC:Immunohistochemistry;LEF:Lymphoidenhancerfactor;PCR:Polymerasechainreaction;PIN1:Peptidyl-prolylcis/transisomerase;TCF:T-cellfactor.
FatimaSetal.
DKKsandWnt/β-cateninsignallingJiangetal[87]reportingmutationsin2of32HCCcases,whereasCuietal[25]didnotreportanymutationin34HCCcases.
However,overexpressionofTCF-4inHCCtissueshasbeencorrelatedtoc-Mycoverexpression,tu-mourcapsulestatusandintrahepaticmetastasis[87].
LEF-1isalsooverexpressedinHCCandassociatedwithcyclinD1overexpression,butitisnotfoundtobecorrelatedwithanyclinicopathologicalparameters[88].
Therefore,itisnotknownwhethertheaberrantexpressionofTCF-4andLEF-1isaprimaryeventinHCCoraresultofab-errantWnt/β-cateninsignallinginHCC.
ExpressionofotherTCF/LEFfamilymembersinHCChasnotbeenreported.
Cadherin-17Cadherin-17(CDH17)belongstothecadherinsuperfam-ilycomprisingtransmembraneglycoproteinsthatmediatecalcium-dependentcell-celladhesion[89].
Sinceβ-cateninplaysanessentialroleincadherin-mediatedcell-celladhe-sionaswellasinWnt/β-cateninsignalling,ourlaboratoryidentifiedCDH17toaffectWnt/β-cateninsignallinginHCC.
CDH17isoverexpressedinHCCcellsandinmorethan70%ofHCCcases[90].
Inaddition,overexpressionofanalternativesplicevariantofCDH17(lackingexon7)inmorethan50%ofHCCiscorrelatedtotumourrecur-rence,venousinfiltrationandreducedoverallsurvival[91].
Interestingly,theCDH17splicevariantisfoundtobetriggeredbytwosinglenucleotidepolymorphismsthatareidentifiedasgeneticfactorscontributingtothehighdevel-opmentofHCCinAsia[92].
Furthermore,RNAinterfer-ence(RNAi)-mediatedknockdownofCDH17inmeta-staticHCCcellshasbeenshowntoinhibittumourigenesisinvivoandreduceWnt/β-cateninsignallingbydecreasingphospho-GSK3βandcyclinD1.
Thiswasaccompaniedbyre-localisationofβ-catenintothecytoplasm[93].
TetraspaninsTetraspaninsaretransmembraneproteinsknowntoaf-fectawiderangeoffunctionsincludingcell-celladhe-sion,cellgrowthandsuppressionofmetastasis[94].
TherecentinvolvementoftetraspaninsCD9andCD82inanovelmechanismtoantagonizeWnt/β-cateninsignallingbyexosomalreleaseofβ-cateninisanexcitingavenuetoexploreinHCC.
Thisexosomalreleaseofβ-cateninmaybecompromisedincancerswithhighWnt/β-cateninsignalling.
CD9andCD82aresuppressorsofmetastasisandtheirexpressionisreducedinHCCwithportalveininvasionand/orintrahepaticmetastasis[95].
Chairoungduaetal[38]demonstratedWnt/β-cateninsignallinginhibitioninametastaticcelllinefollowingrestorationofCD82expression.
Thus,thesetetraspaninsmaysuppressmetas-tasisbyantagonizingWnt/β-cateninsignallingbytarget-ingβ-cateninforexosomalrelease.
ItwillbeimportanttoinvestigatethecorrelationbetweenCD9andCD82withβ-catenininHCC.
MicroRNAsMicroRNAs(miRNAs)aresmallnon-codingRNAsthatregulatepost-transcriptionalgeneexpression[96].
TheyareaberrantlyexpressedinHCCcomparedtotheirnon-tumourlivertissues[97-99]andcontributetolivertumou-rigenesis[100,101].
SeveralmiRNAshavebeenidentifiedtoaffecttheWnt/β-cateninpathway[102].
Usingaglobalmicroarray-basedmiRNAprofilingapproach,Jietal[103]identifiedmiRNA-181(miR-181)tobeupregulatedinHCCtumoursthatwerepositiveforepithelialcelladhe-sionmolecule(EpCAM)andAFP(EpCAM+AFP+).
SuchtumoursdemonstratedcancerstemcellpropertiesandanactivationofWnt/β-cateninsignalling.
InvitrostudiesshowedacorrelationbetweenoverexpressionofmiR-181andβ-catenininHCCcellsandfurtherdemonstratedthatmiR-181promotedthestemnessofEpCAM+AFP+HCCcellsbytargetingCDX2(caudaltypehomeoboxtranscriptionfactor2),GATA6(GATAbindingprotein6,ahepatictranscriptionalregulatorofdifferentiation)andnemo-likekinase(NLK,aninhibitorofWnt/β-cateninsignalling).
ThesefindingsprovideevidencethatmiR-181istranscriptionallyactivatedbyWnt/β-cateninsignallingandinturninhibitsitsregulators.
Inaddition,miR-375isanothermiRNAinvolvedintheWnt/β-cateninpathwayanditisdownregulatedbyβ-catenininHCC[104].
How-ever,thefunctionofmiR-375andthemechanismsbywhichitisregulatedbyβ-cateninarenotclear.
FurtherresearchisneededtoinvestigatetheinvolvementofmiR-NAsinWnt/β-cateninsignallinginHCC.
Yes-associatedproteinTheHipposignallingpathwaycontrolsorgansizebyregulatingcellproliferationandapoptosis.
Thesignallingcascadeofthispathwayultimatelyleadstothephosphor-ylationofyes-associatedprotein(YAP),adownstreameffectorofthispathway.
YAPisatranscriptionalco-ac-tivatoranditsphosphorylationcausesittoremaininthecytoplasmandpreventthetranscriptionofgenesrespon-sibleforcellproliferationandinhibitionofapoptosis[105].
Recently,afewstudieshavedescribedtheHippopathwayasanegativeregulatorofWnt/β-cateninsignalling[106,107].
Varelasetal[106]reportedphosphorylatedTaz(componentoftheHippopathway)toinhibittheactivationofDvl,therebypreventingβ-cateninstabilisationandactivation.
Heallenetal[107]recentlyshowednuclearinteractionofunphosphorylatedYAPandβ-cateninincardiaccellsofmicewithdysregulatedHipposignalling.
ThesemicehadenlargedheartsandoverexpressedWnt/β-catenintargetgenes.
Additionally,dysregulationoftheHipposignallingpathwayandinhibitionofβ-cateninresultedinrestric-tionofcardiomyocyteovergrowth.
Thisstudyofferedinsightsintothedirectinteractionbetweenthedown-streameffectorsofthesetwoimportantpathways.
InHCC,nuclearoverexpressionofYAP(unphosphorylatedYAP)hasbeenreportedin62%ofHCCandhasbeenassociatedwithshortdisease-freesurvivalandoverallsurvival[108].
Sinceβ-cateninisalsofoundtoaccumulateinthenucleusofHCCpatients[43,45],furtherstudiesarewarrantedtounderstandtheclinicalimplicationsofYAPandβ-cateninoverexpressioninHCC.
316August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comFatimaSetal.
DKKsandWnt/β-cateninsignalling317August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comWNTSIGNALLINGANTAGONISTSAsmentionedabove,therearemanyfactorsaffectingtheWnt/β-cateninpathwayinHCC.
However,theoccur-renceofthesefactorsisnotabsoluteineachclinicalcaseofHCC.
Therefore,theinvolvementofotherfactorsinmodulatingtheWnt/β-cateninpathwayisspeculated.
FactorsupstreamofGSK3β/Axin/APCproteincom-plex,likethoseoccurringattheextracellularlevel,mayalsobethoseinvolved.
Forthis,secretedWntantagonistsaredistinguishedexamples.
TheseantagonistscanbedividedbroadlyintotwogroupsbasedontheirdifferentmechanismsinpreventingtheinteractionofWntwithLRP5/LRP6[109].
GroupⅠconsistsofWise,sclerostinanddickkopfs(DKKs)thatbinddirectlytoLRP5/LRP6.
GroupⅡcomprisesWntinhibitoryfactorandsecretedfrizzled-relatedproteinthatactbydirectlybindingtoWntproteinsandpreventtheformationoftheWntreceptorcomplex.
ThisreviewwillfocusontheroleofDKKsontheWnt/β-cateninpathwayinHCC.
StructureofDKKsTheDKKfamilyconsistsoffourmembers(DKK1toDKK4).
Theyaresecretedglycoproteinsof225-350ami-noacidswithmolecularweightsbetween25and29kDaforDKK1,DKK2andDKK4,and38kDaforDKK3.
DKKscontaintwoconservedcysteine-richdomains,eachofwhichisseparatedbyalinkerregionofvariouslengths[110].
Eachcysteine-richdomaincontainstencys-teineresidues.
Theamino-terminalcysteine-richdomain(Cys-1)isuniquetoeachDKK,whereasthecarboxyl-terminalcysteine-richdomain(Cys-2)ishighlyconservedamongallmembersoftheDKKfamily.
ThepositionofeachcysteineresidueintheCys-2domaincloselyre-semblesproteinsinthecolipasefamily[111].
Becauseoftheroleofcolipasesinlipidhydrolysis[112],thepresenceofthisfeaturesuggeststheabilityofDKKstointeractwithlipidsinregulatingWnt/β-cateninsignalling[111].
AmongallDKKs,DKK3isthemostdivergentmember[113].
Itcontainsanextendedamino-terminaldomainprecedingtheCys-1regionandanextendedcarboxyl-terminaldo-mainfollowingtheCys-2domain.
AllDKKspossesssev-eralpotentialsitesforproteolyticcleavagebyfurin-typeproteases,indicatingthattheymaybesubjectedtopost-translationalmodifications[114].
Figure2AandBillustratethedifferencesbetweendifferentDKKs.
FunctionsofDKKsontheWnt/β-cateninpathwayMembersoftheDKKfamilydiffernotonlyintheirstructuresbutalsointheirmRNAexpressioninHCCandintheirabilitytomodulateWnt/β-cateninsignalling.
Table3summarizesthesedifferencesbetweenmembersoftheDKKfamily.
DKK1isthemoststudiedmemberoftheDKKfamily.
ItwasoriginallyidentifiedasaheadinducerwhenitsmRNAwasinjectedintoXenopusembryos[115].
Experi-mentsinactivatingXenopusDKK1withanti-DKK1anti-bodies[115]andinvolvingDKK1knockoutmice[116]showalackofanteriorheadstructures,highlightingitsimpor-tanceinheadformation.
DKK1inhibitsWnt-inducedstabilisationofβ-cateninandtwomodelshavebeenpro-posed.
ThefirstmodelproposesthatDKK1bindstotheextracellulardomainofLRP5/LRP6andpreventstheformationoftheFZD-Wnt-LRP5/LRP6complexinre-sponsetoWnt,therebyattenuatingWntactivity[117].
ThesecondmodelproposesthatDKK1inhibitsWntsignal-lingbyinducingclathrin-dependentinternalisationofLRP6[37].
However,contrarytotheresultsofYamamotoetal[37],Blitzeretal[118]usedmousefibroblastcellstoshowthatclathrin-dependentinternalisationofLRP6isrequiredtopropagateWnt/β-cateninsignalling,anddisturbingthisclathrin-mediatedendocytosisblocksWntactivity.
TheserecentfindingssuggestthatmechanismsofDKK1an-tagonisticactivitymayvaryindifferentcelltypes.
Figure2Domainstructureandphylogenetictreeofhumandickkopfproteins.
A:Eachdickkopf(DKK)containstwocysteine-richdomains,eachofwhichisseparatedbyalinkerregionofvariouslengths.
Theamino-terminalcysteine-richdomain(Cys-1)domainsareuniquetoeachDKK,whereasthecarboxyl-terminalcysteine-richdomain(Cys-2)domainsareconservedamongallmembersoftheDKKfamily;B:PhylogenetictreeoftheDKKproteins.
AminoacidsequenceswerealignedbytheClustalWprogram,andthephylogenetictreewasconstructedbytheneighbourjoiningmethod.
Thescalebarindicatestheestimatednumberofsub-stitutionsper50aminoacids.
SP:Signalpeptide.
DKK30.
05DKK4DKK2DKK1DKK1DKK2DKK3DKK4266aminoacids259aminoacids350aminoacids224aminoacidsScalebar50aminoacidsBALinkerregionSPSPSPSPCys-1Cys-1Cys-1Cys-1Cys-2Cys-2Cys-2Cys-2FatimaSetal.
DKKsandWnt/β-cateninsignalling318August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comInadditiontobindingtoLRP5/LRP6,DKK1alsobindstoKremen1(Krm1)andKrm2,whichbelongtoanotherclassoftransmembranereceptors[119,120].
DKK1bindstoKrm1andKrm2withhighaffinityandthisinhibitsWnt/β-cateninsignalling[121].
Despitethat,geneknockoutstudiesinmicehaveshownthatKrm1andKrm2arenotuniversallyrequiredforDKK1-associatedfunction[122].
ItwasdemonstratedthatDKK1mutantsareabletoantagonizeWntactivitywithoutbindingtoKrm1orKrm2[123].
Therefore,KrmsmaynotbeessentialforDKK1functionandfurtherstudiesareneededtounder-standtheirinvolvementinWnt/β-cateninsignalling.
AmongallDKKs,DKK4demonstratessimilaran-tagonisticactivitytowardstheWnt/β-cateninpathwayasDKK1bybindingtoLRP5/LRP6andKrms[120].
WhilefunctioningupstreamoftheWnt/β-cateninpathway,DKK1andDKK4arealsodownstreamtargetsoftheWnt/β-cateninpathway,creatinganegativefeedbacklooptoregulateWnt/β-cateninsignalling[124,125].
However,thisfeedbackmechanismisoftenabrogatedincancersincludingHCC[125,126].
LikeDKK1andDKK4,DKK2alsobindstoLRP5/LRP6andKrms.
However,DKK2mayserveasanan-tagonistoranagonisttotheWnt/β-cateninpathwaydependingoncellularcontext.
Forinstance,overexpres-sionofDKK2in293fibroblastcellsresultsinWnt/β-cateninpathwayactivation,whereasco-transfectionofDKK2andKrm2inthesamecelltypeinhibitsthispathway[127].
Toexplainthisphenomenon,Chenetal[128]identifiedYWTDβ-propellerdomainsofLRP5/LRP6asthedockingsitesforCys-2ofmouseDKK2,whilethisCys-2domainalsocontainsbindingsiteforKrm1andKrm2.
Therefore,theexpressionofKrmsservesasaswitchforthedualroleofDKK2ontheWnt/β-cateninpathway.
UnlikeotherDKKshavingaroleinWnt/β-cateninsignallingbybindingtoLRP5/LRP6andKrms,thereceptorforDKK3hasnotbeenidentifiedanditsef-fectonthispathwayremainsunclear.
EarlierstudiesinXenopussuggestedthatDKK3isnotinvolvedinWnt/β-cateninsignalling[113,115],butrecentstudieshavedemon-stratedthatDKK3isassociatedwithareductionincyto-plasmicandnuclearaccumulationofβ-catenininSaos-2osteosarcomacells,lungcancercellsandcervicalcancercells[129-131].
Interestingly,Leeetal[130]identifiedβ-TrCPasabindingpartnertoDKK3,andpossiblyDKK3mayreduceβ-cateninlevelsviaubiquitination.
Therefore,thepotentialroleofDKK3inWnt/β-cateninpathwayre-mainstobedetermined.
SincedifferentDKKsexhibitdifferenteffectsonWnt/β-cateninactivity,itwillbeimportanttostudythemechanismsbywhicheachmemberoftheDKKfamilyexertsitseffectontheWnt/β-cateninpathway.
Further-more,asreportedbyYamamotoetal[37]andBlitzeretal[118],DKKsmayhavedifferentmechanismsofactionindif-ferentcelltypes.
Additionally,sharingthesamereceptormaynotimplyexhibitingthesameeffectasinthecaseofDKK1andDKK2.
Forinstance,theybothbindtoLRP5/LRP6andKrms,butDKK1servesasanantago-nist,whereasDKK2servesasbothanagonistandanantagonisttotheWnt/β-cateninpathway.
DKKsinHCCAsmentionedabove,theWnt/β-cateninpathwayplaysacriticalroleinHCCandnotsurprisingly,Wntinhibitors,includingDKKs,areinvolved.
MoreresearchstudieshavebeendedicatedtostudyingtheroleofDKKsinHCCoverthepastfewyears.
DKK1SeveralstudieshavereportedoverexpressionofDKK1inHCCcelllinesandtissues[126,132,133],whileYuetal[126]wasthefirsttodemonstrateacorrelationbetweenDKK1overexpressionandcytoplasmic/nuclearβ-cateninaccu-mulationinHCC.
Itwasdemonstratedthroughinvitroas-saysthatDKK1failedtoinhibitTCF-mediatedtranscrip-tionalactivityinHCCcellswithcytoplasmicornuclearβ-cateninaccumulation,suggestinganabrogationofthenegativefeedbackloopofDKK1inHCC.
SurvivalanalysiscorrelatedoverexpressionofDKK1withpoorprognosisofHCCpatients,andDKK1wasidentifiedasanindependentprognosticmarkerforoverallsurvivalandCys-1:Amino-terminalcysteine-richdomain;Cys-2:Carboxyl-terminalcysteine-richdomain;HCC:Hepatocellularcarcinoma;LRP:Lipoproteinreceptor-relatedprotein;DKK:Dickkopf.
Table3DifferencesbetweentheDKKfamilymembersPropertyDKK1DKK2DKK3DKK4Ref.
VariantsUnknownUnknown2Unknown[130]LocationofCys-185-138aminoacids78-127aminoacids147-195aminoacids41-90aminoacids[110]LocationofCys-2189-263aminoacids183-256aminoacids208-284aminoacids145-218aminoacids[110]Lengthoflinkerregion(numberofaminoacids)50561254[110]ReceptorLRP5/LRP6LRP5/LRP6NotidentifiedLRP5/LRP6[113]ActivityinWnt/β-cateninsignallinginHCCAntagonistNotreportedNotreportedNotreported[126]ExpressioninHCCtissuesOverexpressedReducedReducedNotreported[126,143]PhenotypeofknockoutmiceEmbryoniclethalityViable,fertile,blindness,osteopaeniaViable,fertile,hyperactive,increasedIgM,naturalkillercellsandhaematocritlevelsNotreported[116,162-164]FatimaSetal.
DKKsandWnt/β-cateninsignalling319August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comdisease-freesurvival.
The5-yearoverallsurvivalanddis-ease-freesurvivalratesforHCCpatientsoverexpressingDKK1(43.
4%and34.
2%,respectively)weresignificantlylowerthanHCCpatientswithreducedDKK1expres-sion(59.
3%and55.
2%,respectively).
AlthoughelevatedlevelsofAFPremainthegoldstandardforscreeningHCC,thereare,however,asubgroupofpatientswhohaveHCCandnormallevelsofAFP.
WhenpatientswerestratifiedaccordingtoAFPlevels,DKK1overexpressiondemonstratedworseprognosisforAFP-normalHCCpatients,suggestingthatDKK1mayserveasaprognosticmarkerforthisgroupofpatients.
Furthermore,HCCpatientswithhighDKK1andβ-cateninexpressionalsoshowedpoorprognosis.
The5-yearoverallsurvivalanddisease-freesurvivalrateswere66.
0%and59.
8%,respec-tively,forHCCpatientswithoutDKK1andβ-cateninexpression,and46.
0%and18.
0%forHCCpatientswithhighDKK1andhighβ-cateninexpression[126].
ThisstudyhighlightstheimportantroleofDKK1inWnt/β-cateninsignallinginHCC.
OtherthantotalAFPlevels,lensculinarisagglutinin–reactiveAFP(AFP-L3)andprothrombininducedbyvitaminKabsence-Ⅱ(PIVKA-Ⅱ)havebeenreportedastumourmarkersforHCC.
HighserumAFP-L3levelshaverecentlybeenreportedasaprognosticmarkereveninHCCpatientswithlowAFPlevels[134].
Additionally,highserumlevelsofPIVKA-ⅡhavebeenassociatedwithadvancedHCCwithportalveininvasion[135].
BecauseofitshighexpressioninHCCtissuesanditssecretorynature,DKK1ishypothesizedtobepresentathighlevelsintheserumofHCCpatients.
However,therearenostudiesevaluatinghighDKK1serumlevelsonHCCprogressionorprognosisanditwouldbeim-portanttoconductsuchastudytounderstandtheclinicalsignificanceofDKK1inHCC.
Ontheotherhand,highlevelsofDKK1inpatients'serumareassociatedwithpoorprognosisinvariouscancersincludingoesophagealsquamouscellcarcinoma[136],lungcancer[137],breastcan-cer[138]andcervicalcancer[138],suggestingthattheserumlevelofDKK1mayalsoreflecttheprognosisofHCCpatients.
Inmultiplemyeloma,highDKK1serumlevelsareassociatedwithosteolyticbonelesions[139]andpatientsrespondingtoanti-myelomatreatmentshowadecreaseinDKK1serumlevels[140],suggestingtheinvolvementofDKK1inthisaspect.
Recently,Fulcinitietal[141]evaluatedtheeffectofanti-DKK1monoclonalantibody(BHQ880)inamultiplemyelomamousemodelandfoundthatitinducedboneformationandinhibitedtumour-inducedosteolyticbonelesions.
Likemultiplemyeloma,HCCisalsoosteolyticinnaturewith20%ofHCCpatientshav-ingbonemetastasis[142],makingitimportanttoassesstheroleofDKK1inbonemetastasisinHCC.
DKK2InHCC,ahigherlevelofDKK2methylationwasde-tectedinHCCtissuescomparedtocorrespondingnon-cancerouscirrhotictissues[143],suggestingitsroleinhepatocarcinogenesis.
EpigeneticsilencingofDKK2hasalsobeenreportedingastriccancer[144],oesophagealsquamouscellcarcinoma[145]andrenalcellcarcinoma[146].
Inrenalcellcarcinoma,nosignificantrelationshipwasfoundbetweenDKK2methylationandβ-cateninexpres-sion[146].
AlthoughtheeffectofDKK2onWnt/β-cateninsignallingdependsontheexpressionofLRP5/LRP6andKrms,DKK2expressionhasnotbeenstudiedinthecontextofthesemolecules.
Furthermore,therearecurrentlynoreportsontheeffectofDKK2onWnt/β-cateninsignallinginHCC.
DKK3TherearefewreportsontheclinicalsignificanceofDKK3inHCC[143,147].
ReductioninDKK3expressionisassociatedwithincreasedfrequencyofmethylationinHCCtissuescomparedtocorrespondingnon-cancerouscirrhotictissues,implyingthatDKK3methylationmaynotbeanearlyeventinHCC,butmayfunctionintheprogressionofHCC.
Furthermore,DKK3methylationhasbeensignificantlyassociatedwithshortprogression-freesurvivalinHCCpatients[143].
TheeffectofDKK3onWnt/β-cateninsignallinghasnotbeenreportedinHCC,butreducedDKK3expressionhasbeenshowntocorrelatewithβ-cateninaccumulationinlungcancer[131].
AlthoughtherearenoreportsofDKK3levelinHCCserum,reducedDKK3serumlevelsinovariancancerareassociatedwiththepresenceoflymphnodemetastasis,andhighDKK3serumlevelshavebeenassociatedwithlargetumoursincervicalcancer[148].
Interestingly,mul-tigenemethylationstatusofacombinationofWntan-tagonistgenes,includingDKK3andothers,intheserumhavebeenproposedasmarkersfordiagnosis,stagingandprognosisofrenalcellcarcinoma[149].
AlthoughdifferentWntantagonistsmayfunctiondifferently,theircumula-tivesilencingmayserveasamolecularmarkerforcancerdetection.
DKK4DKK4isleaststudiedincancer.
Itislocatedonchromo-some8p11.
2-p11.
1andthischromosomalregionexperi-encesfrequentlossofheterozygosityinHCC[150].
ThismayexplainreducedexpressionofDKK4inHCCcelllineswithoutdetectionofDKK4methylation[144].
Cur-rentlytherearenoreportsonDKK4expressioninHCCtissuesoritseffectonWnt/β-cateninsignallinginHCC.
Recently,Hirataetal[151]reportedareductionofWnttargetgenesafterectopicexpressionofDKK4inrenalcarcinoma-derivedCakicells.
However,overexpressionofDKK4alsoresultedinactivationoftheJNKpathwayandenhancedtumourgrowthinvivo.
ThissuggeststhatDKK4maybeinvolvedinpathwaysotherthanWnt/β-cateninsignalling.
POTENTIALTHERAPEUTICTARGETSFORWNT/β-CATENINSIGNALLINGINHCCMountingevidencesuggeststheroleofβ-cateninstabi-FatimaSetal.
DKKsandWnt/β-cateninsignalling320August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comlisationinpromotingtumourproliferationinHCCpa-tients[43,152].
SuchobservationsmakeWnt/β-cateninsig-nallinganattractivetargetforcancertherapy.
Inlinewiththishypothesis,smallmoleculeinhibitorshavebeende-velopedtohinderWnt/β-cateninsignallingbydisruptingprotein-proteininteractionsofcomponentsofthispath-way.
Theseincludethefungalderivatives,PKF115-584andCGP049090,thatfunctiontoimpedetheinteractionbetweenβ-cateninandtheTCFcomplex[153].
Emamietal[154]reportedanothermoleculecalledICG-001,whichdisruptstheinteractionbetweenβ-cateninandCBP.
Morerecently,pyrviniumpamoate,ananthelminticdrug,wasshowntoinhibittheWnt/β-cateninpathwaybyallo-stericactivationofCKIαandsubsequentlyincreasedlev-elsofβ-catenindestructioncomplexsecondarytoAxinstabilisation[155].
TherapeuticantibodiesagainstWnt1andWnt2havealsodemonstratedWnt/β-cateninsignallinginhibitionandsuppressionoftumourgrowthinvivo[156,157].
β-cateninsuppressionthroughchemoprophylaxismayofferanotheralternativetherapy.
R-Etodolac(enantiomerofthenon-steroidalanti-inflammatorydrugEtodolac)reducedproliferationandsurvivaloftwoHCCcelllines(HepG2andHep3B)bydecreasingthetotalandactiveforms(dephosphorylatedatSer33/Ser37/Thr41)ofβ-catenin[158].
FurtherstudieswillbeimportanttoaccessclinicalimplicationsofthesepotentialtargetsinHCC.
PERSPECTIVESAberrantactivationofWnt/β-cateninsignallingisanimportanteventinHCCthatleadstotranscriptionofvarioustargetgenesinvolvedincarcinogenesis.
ItwillbeclinicallyrelevanttotargettheWnt/β-cateninpathwayandregulateitsactivity.
However,sincethepathwaycanbeaberrantlyactivatedbythedysfunctionofseveralgenes,itisimportanttoensurethatwhenonememberistargetedtheotherfunctionsofthesametarget,ifany,arenotdisrupted.
Forinstance,β-cateninisinvolvedintheWnt/β-cateninpathwayandplaysacriticalroleincell-celladhesion.
Therefore,careshouldbetakenindesigningreagentsfordisruptionofmoleculeswithdualfunctions.
Fortumourswithmutatedβ-catenin,oneapproachcouldbetoutilizeRNAitargetingmutantβ-cateninfortherapy,asthiswilltargetselectivelyfortumourcells,butnothealthycells.
However,mutantβ-catenin-negativetumourswithaberrantWnt/β-cateninsignallingofferthegreatestchallengeastherecouldbeamultitudeofreasonsforaberrantWnt/β-cateninactiva-tion.
Therefore,alteringWnt/β-cateninsignallingmaybeadvantageousfortherapeuticmeansbutthereareca-veatsandlimitationsinvolved.
Inlinewiththeseinvestigations,DKKshavebeendiscoveredoverthepast12yearsfortheirabilitiestoin-terfereintheWnt/β-cateninpathway.
Sincethen,muchefforthasbeendedicatedtoexploringtheirfunctionsandimplicationsinliverdevelopmentanddisease.
Dif-ferentDKKsexhibitdifferentexpressionsandfunctionsinvariouscancers.
Someimportantquestionsremaintobeanswered.
Forexample,whereareDKKslocalisedinHCCcellsIstheirexpressionassociatedwithcytoplas-mic/nuclearβ-cateninaccumulationinHCCcellsDoDKKshaveprognosticvalueinHCCpatientsDotheyexerttheireffectsonbothwildtypeandmutantβ-catenininHCCcellsDoDKKsdegradeβ-cateninviaaubiq-uitination-dependentmechanismAredifferentDKKsinvolvedindifferentstagesofHCCdevelopmentWhatisthereceptorforDKK3AreDKKsredundantintheirfunctionsFurthermore,thenegativefeedbackmecha-nismofDKK1alsowarrantsfurtherresearch.
AlthoughDKK1inhibitsWnt/β-cateninsignalling,itsoverexpres-sionhasnoeffectonHCCcellswithcytoplasmic/nuclearβ-cateninaccumulation[126].
ThismaybesuggestiveofanabrogatednegativefeedbacklooporitmayalsoimplythattheinhibitoryeffectofDKK1isonlyfunctionalun-tilreachingapointofsaturationbeyondwhichitcannotexertitsinhibitoryeffect.
Insummary,comparedtotheoverwhelmingbodyofevidenceshowingthefunctionalroleofWnt/β-cateninsignallinginHCC,thebiologicalandphysiologicalrolesofDKKsinrelationtothispath-wayremaintobediscovered.
DKKsprovideanewav-enuetoexploreandtheyaddanotherlevelofcomplexitytothealreadycomplexWnt/β-cateninsignallinginHCC.
REFERENCES1JemalA,BrayF,CenterMM,FerlayJ,WardE,FormanD.
Globalcancerstatistics.
CACancerJClin2011;61:69-902WandsJ.
Hepatocellularcarcinomaandsex.
NEnglJMed2007;357:1974-19763AltekruseSF,McGlynnKA,ReichmanME.
Hepatocellu-larcarcinomaincidence,mortality,andsurvivaltrendsintheUnitedStatesfrom1975to2005.
JClinOncol2009;27:1485-14914AschaMS,HanounehIA,LopezR,TamimiTA,FeldsteinAF,ZeinNN.
Theincidenceandriskfactorsofhepatocel-lularcarcinomainpatientswithnonalcoholicsteatohepatitis.
Hepatology2010;51:1972-19785SeveriT,vanMalensteinH,VerslypeC,vanPeltJF.
Tumorinitiationandprogressioninhepatocellularcarcinoma:riskfactors,classification,andtherapeutictargets.
ActaPharmacolSin2010;31:1409-14206KanwalF,HoangT,KramerJR,AschSM,GoetzMB,Zeri-ngueA,RichardsonP,El-SeragHB.
IncreasingprevalenceofHCCandcirrhosisinpatientswithchronichepatitisCvirusinfection.
Gastroenterology2011;140:1182-1188.
e17NguyenVT,LawMG,DoreGJ.
HepatitisB-relatedhepato-cellularcarcinoma:epidemiologicalcharacteristicsanddis-easeburden.
JViralHepat2009;16:453-4638KudoM,HanKH,KokudoN,ChengAL,ChoiBI,FuruseJ,IzumiN,ParkJW,PoonRT,SakamotoM.
LiverCancerWorkingGroupreport.
JpnJClinOncol2010;40Suppl1:i19-i279MazzaferroV,ChunYS,PoonRT,SchwartzME,YaoFY,MarshJW,BhooriS,LeeSG.
Livertransplantationforhepa-tocellularcarcinoma.
AnnSurgOncol2008;15:1001-100710KimH,RhimH,ChoiD,LimHK,KimYS,LeeWJ,JohJW.
Recurrenceandtreatmentpatterninlong-termsurvivorswithhepatocellularcarcinoma:acomparisonbetweenra-diofrequencyablationandsurgeryasafirst-linetreatment.
WorldJSurg2010;34:1881-188611VivarelliM,GuglielmiA,RuzzenenteA,CucchettiA,Bel-lusciR,CordianoC,CavallariA.
SurgicalresectionversuspercutaneousradiofrequencyablationinthetreatmentofFatimaSetal.
DKKsandWnt/β-cateninsignallingFatimaSetal.
DKKsandWnt/β-cateninsignalling321August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comhepatocellularcarcinomaoncirrhoticliver.
AnnSurg2004;240:102-10712UenoS,SakodaM,KuboF,HiwatashiK,TatenoT,BabaY,HasegawaS,TsubouchiH.
Surgicalresectionversusra-diofrequencyablationforsmallhepatocellularcarcinomaswithintheMilancriteria.
JHepatobiliaryPancreatSurg2009;16:359-36613SimoKA,SereikaSE,NewtonKN,GerberDA.
Laparoscop-ic-assistedmicrowaveablationforhepatocellularcarcinoma:Safetyandefficacyincomparisonwithradiofrequencyabla-tion.
JSurgOncol2011;Epubaheadofprint14HuppertP.
Currentconceptsintransarterialchemoembo-lizationofhepatocellularcarcinoma.
AbdomImaging2011;Epubaheadofprint15LencioniR.
Loco-regionaltreatmentofhepatocellularcarci-noma.
Hepatology2010;52:762-77316CajaL,SanchoP,BertranE,Iglesias-SerretD,GilJ,FabregatI.
OveractivationoftheMEK/ERKpathwayinlivertumorcellsconfersresistancetoTGF-{beta}-inducedcelldeaththroughimpairingup-regulationoftheNADPHoxidaseNOX4.
CancerRes2009;69:7595-760217CalvisiDF,WangC,HoC,LaduS,LeeSA,MattuS,Deste-fanisG,DeloguS,ZimmermannA,EricssonJ,BrozzettiS,StanisciaT,ChenX,DombrowskiF,EvertM.
Increasedlipogenesis,inducedbyAKT-mTORC1-RPS6signaling,promotesdevelopmentofhumanhepatocellularcarcinoma.
Gastroenterology2011;140:1071-108318ChangQ,ZhangY,BeezholdKJ,BhatiaD,ZhaoH,ChenJ,CastranovaV,ShiX,ChenF.
SustainedJNK1activationisassociatedwithalteredhistoneH3methylationsinhumanlivercancer.
JHepatol2009;50:323-33319HeY,ZhangH,YinJ,XieJ,TanX,LiuS,ZhangQ,LiC,ZhaoJ,WangH,CaoG.
IkappaBalphagenepromoterpoly-morphismsareassociatedwithhepatocarcinogenesisinpatientsinfectedwithhepatitisBvirusgenotypeC.
Carcino-genesis2009;30:1916-192220WhittakerS,MaraisR,ZhuAX.
Theroleofsignalingpath-waysinthedevelopmentandtreatmentofhepatocellularcarcinoma.
Oncogene2010;29:4989-500521ApteU,ThompsonMD,CuiS,LiuB,CieplyB,MongaSP.
Wnt/beta-cateninsignalingmediatesovalcellresponseinrodents.
Hepatology2008;47:288-29522Nejak-BowenKN,MongaSP.
Beta-cateninsignaling,liverregenerationandhepatocellularcancer:sortingthegoodfromthebad.
SeminCancerBiol2011;21:44-5823GonzalezFJ.
Roleofbeta-cateninintheadultliver.
Hepatol-ogy2006;43:650-65324HsiehA,KimHS,LimSO,YuDY,JungG.
HepatitisBviralXproteininteractswithtumorsuppressoradenomatouspol-yposiscolitoactivateWnt/β-cateninsignaling.
CancerLett2011;300:162-17225CuiJ,ZhouX,LiuY,TangZ,RomeihM.
Wntsignalinginhepatocellularcarcinoma:analysisofmutationandexpres-sionofbeta-catenin,T-cellfactor-4andglycogensynthasekinase3-betagenes.
JGastroenterolHepatol2003;18:280-28726CalvisiDF,FactorVM,LaduS,ConnerEA,ThorgeirssonSS.
Disruptionofbeta-cateninpathwayorgenomicinstabil-itydefinetwodistinctcategoriesoflivercancerintransgenicmice.
Gastroenterology2004;126:1374-138627KemlerR.
Fromcadherinstocatenins:cytoplasmicproteininteractionsandregulationofcelladhesion.
TrendsGenet1993;9:317-32128DreesF,PokuttaS,YamadaS,NelsonWJ,WeisWI.
Alpha-cateninisamolecularswitchthatbindsE-cadherin-beta-cateninandregulatesactin-filamentassembly.
Cell2005;123:903-91529SakanakaC.
Phosphorylationandregulationofbeta-cateninbycaseinkinaseIepsilon.
JBiochem2002;132:697-70330OrfordK,CrockettC,JensenJP,WeissmanAM,ByersSW.
Serinephosphorylation-regulatedubiquitinationanddegra-dationofbeta-catenin.
JBiolChem1997;272:24735-2473831MaherMT,MoR,FlozakAS,PeledON,GottardiCJ.
Beta-cateninphosphorylatedatserine45isspatiallyuncoupledfrombeta-cateninphosphorylatedintheGSK3domain:im-plicationsforsignaling.
PLoSOne2010;5:e1018432ZengX,HuangH,TamaiK,ZhangX,HaradaY,YokotaC,AlmeidaK,WangJ,DobleB,WoodgettJ,Wynshaw-BorisA,HsiehJC,HeX.
InitiationofWntsignaling:controlofWntcoreceptorLrp6phosphorylation/activationviafrizzled,di-shevelledandaxinfunctions.
Development2008;135:367-37533ZengX,TamaiK,DobleB,LiS,HuangH,HabasR,Okamu-raH,WoodgettJ,HeX.
Adual-kinasemechanismforWntco-receptorphosphorylationandactivation.
Nature2005;438:873-87734BrantjesH,BarkerN,vanEsJ,CleversH.
TCF:LadyJusticecastingthefinalverdictontheoutcomeofWntsignalling.
BiolChem2002;383:255-26135DanielsDL,WeisWI.
Beta-catenindirectlydisplacesGroucho/TLErepressorsfromTcf/LefinWnt-mediatedtranscriptionactivation.
NatStructMolBiol2005;12:364-37136YamamotoH,KomekadoH,KikuchiA.
Caveolinisneces-saryforWnt-3a-dependentinternalizationofLRP6andac-cumulationofbeta-catenin.
DevCell2006;11:213-22337YamamotoH,SakaneH,YamamotoH,MichiueT,KikuchiA.
Wnt3aandDkk1regulatedistinctinternalizationpathwaysofLRP6totunetheactivationofbeta-cateninsignaling.
DevCell2008;15:37-4838ChairoungduaA,SmithDL,PochardP,HullM,CaplanMJ.
Exosomereleaseofβ-catenin:anovelmechanismthatan-tagonizesWntsignaling.
JCellBiol2010;190:1079-109139LotvallJ,ValadiH.
CelltocellsignallingviaexosomesthroughesRNA.
CellAdhMigr2007;1:156-15840SuzukiT,YanoH,NakashimaY,NakashimaO,KojiroM.
Beta-cateninexpressioninhepatocellularcarcinoma:apos-sibleparticipationofbeta-catenininthededifferentiationprocess.
JGastroenterolHepatol2002;17:994-100041WongCM,FanST,NgIO.
beta-Cateninmutationandover-expressioninhepatocellularcarcinoma:clinicopathologicandprognosticsignificance.
Cancer2001;92:136-14542HuangH,FujiiH,SankilaA,Mahler-AraujoBM,MatsudaM,CathomasG,OhgakiH.
Beta-cateninmutationsarefrequentinhumanhepatocellularcarcinomasassociatedwithhepati-tisCvirusinfection.
AmJPathol1999;155:1795-180143NhieuJT,RenardCA,WeiY,CherquiD,ZafraniES,Buen-diaMA.
Nuclearaccumulationofmutatedbeta-catenininhepatocellularcarcinomaisassociatedwithincreasedcellproliferation.
AmJPathol1999;155:703-71044MaoTL,ChuJS,JengYM,LaiPL,HsuHC.
Expressionofmutantnuclearbeta-catenincorrelateswithnon-invasivehepatocellularcarcinoma,absenceofportalveinspread,andgoodprognosis.
JPathol2001;193:95-10145InagawaS,ItabashiM,AdachiS,KawamotoT,HoriM,ShimazakiJ,YoshimiF,FukaoK.
Expressionandprognosticrolesofbeta-catenininhepatocellularcarcinoma:correlationwithtumorprogressionandpostoperativesurvival.
ClinCancerRes2002;8:450-45646KondoY,KanaiY,SakamotoM,GendaT,MizokamiM,UedaR,HirohashiS.
Beta-cateninaccumulationandmuta-tionofexon3ofthebeta-cateningeneinhepatocellularcar-cinoma.
JpnJCancerRes1999;90:1301-130947CieplyB,ZengG,Proverbs-SinghT,GellerDA,MongaSP.
Uniquephenotypeofhepatocellularcancerswithexon-3mutationsinbeta-cateningene.
Hepatology2009;49:821-83148Laurent-PuigP,LegoixP,BluteauO,BelghitiJ,FrancoD,Bi-notF,MongesG,ThomasG,Bioulac-SageP,Zucman-RossiJ.
Geneticalterationsassociatedwithhepatocellularcarcino-masdefinedistinctpathwaysofhepatocarcinogenesis.
Gas-troenterology2001;120:1763-177349CadoretA,OvejeroC,Saadi-KheddouciS,SouilE,FabreM,RomagnoloB,KahnA,PerretC.
Hepatomegalyintransgen-FatimaSetal.
DKKsandWnt/β-cateninsignalling322August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comicmiceexpressinganoncogenicformofbeta-catenin.
CancerRes2001;61:3245-324950HaradaN,MiyoshiH,MuraiN,OshimaH,TamaiY,Oshi-maM,TaketoMM.
Lackoftumorigenesisinthemouseliverafteradenovirus-mediatedexpressionofadominantstablemutantofbeta-catenin.
CancerRes2002;62:1971-197751TanX,ApteU,MicsenyiA,KotsagrelosE,LuoJH,Ranga-nathanS,MongaDK,BellA,MichalopoulosGK,MongaSP.
Epidermalgrowthfactorreceptor:anoveltargetoftheWnt/beta-cateninpathwayinliver.
Gastroenterology2005;129:285-30252ColnotS,DecaensT,Niwa-KawakitaM,GodardC,HamardG,KahnA,GiovanniniM,PerretC.
Liver-targeteddisrup-tionofApcinmiceactivatesbeta-cateninsignalingandleadstohepatocellularcarcinomas.
ProcNatlAcadSciUSA2004;101:17216-1722153ProsperiJR,BecherKR,WillsonTA,CollinsMH,WitteDP,GossKH.
TheAPCtumorsuppressorisrequiredforepi-thelialintegrityinthemousemammarygland.
JCellPhysiol2009;220:319-33154QianJ,SarnaikAA,BonneyTM,KeirseyJ,CombsKA,Stei-gerwaldK,AcharyaS,BehbehaniGK,BartonMC,LowyAM,GrodenJ.
TheAPCtumorsuppressorinhibitsDNAreplicationbydirectlybindingtoDNAviaitscarboxylter-minus.
Gastroenterology2008;135:152-16255KawateS,FukusatoT,OhwadaS,WatanukiA,MorishitaY.
Amplificationofc-mycinhepatocellularcarcinoma:cor-relationwithclinicopathologicfeatures,proliferativeactivityandp53overexpression.
Oncology1999;57:157-16356GotohJ,ObataM,YoshieM,KasaiS,OgawaK.
CyclinD1over-expressioncorrelateswithbeta-cateninactivation,butnotwithH-rasmutations,andphosphorylationofAkt,GSK3betaandERK1/2inmousehepaticcarcinogenesis.
Carcinogenesis2003;24:435-44257ChanKL,GuanXY,NgIO.
High-throughputtissuemicroar-rayanalysisofc-mycactivationinchronicliverdiseasesandhepatocellularcarcinoma.
HumPathol2004;35:1324-133158SatoY,ItohF,HareyamaM,SatohM,HinodaY,SetoM,UedaR,ImaiK.
AssociationofcyclinD1expressionwithfactorscorrelatedwithtumorprogressioninhumanhepato-cellularcarcinoma.
JGastroenterol1999;34:486-49359NishidaN,FukudaY,KomedaT,KitaR,SandoT,Furu-kawaM,AmenomoriM,ShibagakiI,NakaoK,IkenagaM.
AmplificationandoverexpressionofthecyclinD1geneinaggressivehumanhepatocellularcarcinoma.
CancerRes1994;54:3107-311060JooM,KangYK,KimMR,LeeHK,JangJJ.
CyclinD1over-expressioninhepatocellularcarcinoma.
Liver2001;21:89-9561UetaT,IkeguchiM,HirookaY,KaibaraN,TeradaT.
Beta-cateninandcyclinD1expressioninhumanhepatocellularcarcinoma.
OncolRep2002;9:1197-120362deLaCosteA,RomagnoloB,BilluartP,RenardCA,Buen-diaMA,SoubraneO,FabreM,ChellyJ,BeldjordC,KahnA,PerretC.
Somaticmutationsofthebeta-cateningenearefre-quentinmouseandhumanhepatocellularcarcinomas.
ProcNatlAcadSciUSA1998;95:8847-885163CadoretA,OvejeroC,TerrisB,SouilE,LévyL,LamersWH,KitajewskiJ,KahnA,PerretC.
Newtargetsofbeta-cateninsignalingintheliverareinvolvedintheglutaminemetabo-lism.
Oncogene2002;21:8293-830164YamamotoY,SakamotoM,FujiiG,TsuijiH,KenetakaK,AsakaM,HirohashiS.
OverexpressionoforphanG-protein-coupledreceptor,Gpr49,inhumanhepatocellularcarci-nomaswithbeta-cateninmutations.
Hepatology2003;37:528-53365PangR,YuenJ,YuenMF,LaiCL,LeeTK,ManK,PoonRT,FanST,WongCM,NgIO,KwongYL,TseE.
PIN1overex-pressionandbeta-cateningenemutationsaredistinctonco-geniceventsinhumanhepatocellularcarcinoma.
Oncogene2004;23:4182-418666NusseR,VarmusHE.
Wntgenes.
Cell1992;69:1073-108767MoonRT,BrownJD,TorresM.
WNTsmodulatecellfateandbehaviorduringvertebratedevelopment.
TrendsGenet1997;13:157-16268KolligsFT,BommerG,GkeB.
Wnt/beta-catenin/tcfsig-naling:acriticalpathwayingastrointestinaltumorigenesis.
Digestion2002;66:131-14469LeeHH,UenYH,TianYF,SunCS,SheuMJ,KuoHT,KoayLB,LinCY,TzengCC,ChengCJ,TangLY,TsaiSL,WangAH.
Wnt-1proteinasaprognosticbiomarkerforhepatitisB-relatedandhepatitisC-relatedhepatocellularcarcinomaaftersurgery.
CancerEpidemiolBiomarkersPrev2009;18:1562-156970KimM,LeeHC,TsedensodnomO,HartleyR,LimYS,YuE,MerleP,WandsJR.
FunctionalinteractionbetweenWnt3andFrizzled-7leadstoactivationoftheWnt/beta-cateninsignalingpathwayinhepatocellularcarcinomacells.
JHepa-tol2008;48:780-79171BengocheaA,deSouzaMM,LefranoisL,LeRouxE,GalyO,CheminI,KimM,WandsJR,TrepoC,HainautP,ScoazecJY,VitvitskiL,MerleP.
CommondysregulationofWnt/Frizzledreceptorelementsinhumanhepatocellularcarci-noma.
BrJCancer2008;99:143-15072MerleP,delaMonteS,KimM,HerrmannM,TanakaS,VonDemBusscheA,KewMC,TrepoC,WandsJR.
Functionalconsequencesoffrizzled-7receptoroverexpressioninhu-manhepatocellularcarcinoma.
Gastroenterology2004;127:1110-112273BanKC,SinghH,KrishnanR,SeowHF.
GSK-3betaphos-phorylationandalterationofbeta-catenininhepatocellularcarcinoma.
CancerLett2003;199:201-20874Desbois-MouthonC,Blivet-VanEggelpolMJ,BeurelE,BoissanM,DeléloR,CadoretA,CapeauJ.
Dysregulationofglycogensynthasekinase-3betasignalinginhepatocellularcarcinomacells.
Hepatology2002;36:1528-153675JhoEH,ZhangT,DomonC,JooCK,FreundJN,CostantiniF.
Wnt/beta-catenin/TcfsignalinginducesthetranscriptionofAxin2,anegativeregulatorofthesignalingpathway.
MolCellBiol2002;22:1172-118376KimYD,ParkCH,KimHS,ChoiSK,RewJS,KimDY,KohYS,JeungKW,LeeKH,LeeJS,JuhngSW,LeeJH.
GeneticalterationsofWntsignalingpathway-associatedgenesinhepatocellularcarcinoma.
JGastroenterolHepatol2008;23:110-11877TaniguchiK,RobertsLR,AdercaIN,DongX,QianC,MurphyLM,NagorneyDM,BurgartLJ,RochePC,SmithDI,RossJA,LiuW.
Mutationalspectrumofbeta-catenin,AXIN1,andAXIN2inhepatocellularcarcinomasandhepa-toblastomas.
Oncogene2002;21:4863-487178SatohS,DaigoY,FurukawaY,KatoT,MiwaN,NishiwakiT,KawasoeT,IshiguroH,FujitaM,TokinoT,SasakiY,ImaokaS,MurataM,ShimanoT,YamaokaY,NakamuraY.
AXIN1mutationsinhepatocellularcarcinomas,andgrowthsup-pressionincancercellsbyvirus-mediatedtransferofAXIN1.
NatGenet2000;24:245-25079Zucman-RossiJ,BenhamoucheS,GodardC,BoyaultS,GrimberG,BalabaudC,CunhaAS,Bioulac-SageP,PerretC.
DifferentialeffectsofinactivatedAxin1andactivatedbeta-cateninmutationsinhumanhepatocellularcarcinomas.
On-cogene2007;26:774-78080UmTH,KimH,OhBK,KimMS,KimKS,JungG,ParkYN.
AberrantCpGislandhypermethylationindysplasticnodulesandearlyHCCofhepatitisBvirus-relatedhumanmultistephepatocarcinogenesis.
JHepatol2011;54:939-94781KishidaS,YamamotoH,HinoS,IkedaS,KishidaM,Kiku-chiA.
DIXdomainsofDvlandaxinarenecessaryforproteininteractionsandtheirabilitytoregulatebeta-cateninstabil-ity.
MolCellBiol1999;19:4414-442282ChanDW,ChanCY,YamJW,ChingYP,NgIO.
Prickle-1negativelyregulatesWnt/beta-cateninpathwaybypromot-FatimaSetal.
DKKsandWnt/β-cateninsignalling323August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comingDishevelledubiquitination/degradationinlivercancer.
Gastroenterology2006;131:1218-122783YauTO,ChanCY,ChanKL,LeeMF,WongCM,FanST,NgIO.
HDPR1,anovelinhibitoroftheWNT/beta-cateninsig-naling,isfrequentlydownregulatedinhepatocellularcarci-noma:involvementofmethylation-mediatedgenesilencing.
Oncogene2005;24:1607-161484RyoA,NakamuraM,WulfG,LiouYC,LuKP.
Pin1regu-latesturnoverandsubcellularlocalizationofbeta-cateninbyinhibitingitsinteractionwithAPC.
NatCellBiol2001;3:793-80185WatermanML.
Lymphoidenhancerfactor/Tcellfactorex-pressionincolorectalcancer.
CancerMetastasisRev2004;23:41-5286VandeWeteringM,CastropJ,KorinekV,CleversH.
Exten-sivealternativesplicinganddualpromoterusagegenerateTcf-1proteinisoformswithdifferentialtranscriptioncontrolproperties.
MolCellBiol1996;16:745-75287JiangY,ZhouXD,LiuYK,WuX,HuangXW.
AssociationofhTcf-4geneexpressionandmutationwithclinicopathologi-calcharacteristicsofhepatocellularcarcinoma.
WorldJGas-troenterol2002;8:804-80788Schmitt-GraeffA,Ertelt-HeitzmannV,AllgaierHP,OlschewskiM,NitschkeR,HaxelmansS,KoelbleK,BehrensJ,BlumHE.
CoordinatedexpressionofcyclinD1andLEF-1/TCFtranscriptionfactorisrestrictedtoasubsetofhepatocel-lularcarcinoma.
LiverInt2005;25:839-84789LeeNP,PoonRT,ShekFH,NgIO,LukJM.
Roleofcad-herin-17inoncogenesisandpotentialtherapeuticimplica-tionsinhepatocellularcarcinoma.
BiochimBiophysActa2010;1806:138-14590WongBW,LukJM,NgIO,HuMY,LiuKD,FanST.
Identi-ficationofliver-intestinecadherininhepatocellularcarcino-ma--apotentialdiseasemarker.
BiochemBiophysResCommun2003;311:618-62491WangXQ,LukJM,LeungPP,WongBW,StanbridgeEJ,FanST.
AlternativemRNAsplicingofliverintestine-cadherininhepatocellularcarcinoma.
ClinCancerRes2005;11:483-48992WangXQ,LukJM,Garcia-BarceloM,MiaoX,LeungPP,HoDW,CheungST,LamBY,CheungCK,WongAS,LauSS,SoMT,YuWC,CaiQ,LiuKS,HuiCK,LauGK,PoonRT,WongJ,FanST.
Liverintestine-cadherin(CDH17)haplotypeisassociatedwithincreasedriskofhepatocellularcarcinoma.
ClinCancerRes2006;12:5248-525293LiuLX,LeeNP,ChanVW,XueW,ZenderL,ZhangC,MaoM,DaiH,WangXL,XuMZ,LeeTK,NgIO,ChenY,KungHF,LoweSW,PoonRT,WangJH,LukJM.
Targetingcadherin-17inactivatesWntsignalingandinhibitstumorgrowthinlivercarcinoma.
Hepatology2009;50:1453-146394ZllerM.
Tetraspanins:pushandpullinsuppressingandpromotingmetastasis.
NatRevCancer2009;9:40-5595KanetakaK,SakamotoM,YamamotoY,YamasakiS,LanzaF,KanematsuT,HirohashiS.
OverexpressionoftetraspaninCO-029inhepatocellularcarcinoma.
JHepatol2001;35:637-64296JiJ,WangXW.
Newkidsontheblock:diagnosticandprog-nosticmicroRNAsinhepatocellularcarcinoma.
CancerBiolTher2009;8:1686-169397BoutzDR,CollinsPJ,SureshU,LuM,RamírezCM,Fernán-dez-HernandoC,HuangY,AbreuRdeS,LeSY,ShapiroBA,LiuAM,LukJM,AldredSF,TrinkleinND,MarcotteEM,PenalvaLO.
Two-tieredapproachidentifiesanetworkofcancerandliverdisease-relatedgenesregulatedbymiR-122.
JBiolChem2011;286:18066-1807898BurchardJ,ZhangC,LiuAM,PoonRT,LeeNP,WongKF,ShamPC,LamBY,FergusonMD,TokiwaG,SmithR,Lee-sonB,BeardR,LambJR,LimL,MaoM,DaiH,LukJM.
mi-croRNA-122asaregulatorofmitochondrialmetabolicgenenetworkinhepatocellularcarcinoma.
MolSystBiol2010;6:40299LiuAM,ZhangC,BurchardJ,FanST,WongKF,DaiH,PoonRT,LukJM.
GlobalregulationonmicroRNAinhepati-tisBvirus-associatedhepatocellularcarcinoma.
OMICS2011;15:187-191100PineauP,VoliniaS,McJunkinK,MarchioA,BattistonC,TerrisB,MazzaferroV,LoweSW,CroceCM,DejeanA.
miR-221overexpressioncontributestolivertumorigenesis.
ProcNatlAcadSciUSA2010;107:264-269101WongCC,WongCM,TungEK,AuSL,LeeJM,PoonRT,ManK,NgIO.
ThemicroRNAmiR-139suppressesmetas-tasisandprogressionofhepatocellularcarcinomabydown-regulatingRho-kinase2.
Gastroenterology2011;140:322-331102HuangK,ZhangJX,HanL,YouYP,JiangT,PuPY,KangCS.
MicroRNArolesinbeta-cateninpathway.
MolCancer2010;9:252103JiJ,YamashitaT,BudhuA,ForguesM,JiaHL,LiC,DengC,WauthierE,ReidLM,YeQH,QinLX,YangW,WangHY,TangZY,CroceCM,WangXW.
IdentificationofmicroR-NA-181bygenome-widescreeningasacriticalplayerinEpCAM-positivehepaticcancerstemcells.
Hepatology2009;50:472-480104LadeiroY,CouchyG,BalabaudC,Bioulac-SageP,PelletierL,RebouissouS,Zucman-RossiJ.
MicroRNAprofilinginhepatocellulartumorsisassociatedwithclinicalfeaturesandoncogene/tumorsuppressorgenemutations.
Hepatology2008;47:1955-1963105LiuAM,XuMZ,ChenJ,PoonRT,LukJM.
TargetingYAPandHipposignalingpathwayinlivercancer.
ExpertOpinTherTargets2010;14:855-868106VarelasX,MillerBW,SopkoR,SongS,GregorieffA,Fel-louseFA,SakumaR,PawsonT,HunzikerW,McNeillH,WranaJL,AttisanoL.
TheHippopathwayregulatesWnt/beta-cateninsignaling.
DevCell2010;18:579-591107HeallenT,ZhangM,WangJ,Bonilla-ClaudioM,KlysikE,JohnsonRL,MartinJF.
HippopathwayinhibitsWntsignal-ingtorestraincardiomyocyteproliferationandheartsize.
Science2011;332:458-461108XuMZ,YaoTJ,LeeNP,NgIO,ChanYT,ZenderL,LoweSW,PoonRT,LukJM.
Yes-associatedproteinisanindepen-dentprognosticmarkerinhepatocellularcarcinoma.
Cancer2009;115:4576-4585109KawanoY,KyptaR.
SecretedantagonistsoftheWntsignal-lingpathway.
JCellSci2003;116:2627-2634110KrupnikVE,SharpJD,JiangC,RobisonK,ChickeringTW,AmaravadiL,BrownDE,GuyotD,MaysG,LeibyK,ChangB,DuongT,GoodearlAD,GearingDP,SokolSY,McCarthySA.
FunctionalandstructuraldiversityofthehumanDick-kopfgenefamily.
Gene1999;238:301-313111AravindL,KooninEV.
Acolipasefoldinthecarboxy-termi-naldomainoftheWntantagonists--theDickkopfs.
CurrBiol1998;8:R477-R478112vanTilbeurghH,BezzineS,CambillauC,VergerR,CarrièreF.
Colipase:structureandinteractionwithpancreaticlipase.
BiochimBiophysActa1999;1441:173-184113NiehrsC.
FunctionandbiologicalrolesoftheDickkopffam-ilyofWntmodulators.
Oncogene2006;25:7469-7481114NakayamaK.
Furin:amammaliansubtilisin/Kex2p-likeendoproteaseinvolvedinprocessingofawidevarietyofprecursorproteins.
BiochemJ1997;327(Pt3):625-635115GlinkaA,WuW,DeliusH,MonaghanAP,BlumenstockC,NiehrsC.
Dickkopf-1isamemberofanewfamilyofse-cretedproteinsandfunctionsinheadinduction.
Nature1998;391:357-362116MukhopadhyayM,ShtromS,Rodriguez-EstebanC,ChenL,TsukuiT,GomerL,DorwardDW,GlinkaA,GrinbergA,HuangSP,NiehrsC,IzpisúaBelmonteJC,WestphalH.
Dickkopf1isrequiredforembryonicheadinductionandlimbmorphogenesisinthemouse.
DevCell2001;1:423-434117SemnovMV,TamaiK,BrottBK,KühlM,SokolS,HeX.
HeadinducerDickkopf-1isaligandforWntcoreceptorFatimaSetal.
DKKsandWnt/β-cateninsignalling324August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comLRP6.
CurrBiol2001;11:951-961118BlitzerJT,NusseR.
AcriticalroleforendocytosisinWntsig-naling.
BMCCellBiol2006;7:28119NakamuraT,NakamuraT,MatsumotoK.
ThefunctionsandpossiblesignificanceofKremenasthegatekeeperofWntsig-nallingindevelopmentandpathology.
JCellMolMed2008;12:391-408120MaoB,WuW,DavidsonG,MarholdJ,LiM,MechlerBM,DeliusH,HoppeD,StannekP,WalterC,GlinkaA,NiehrsC.
KremenproteinsareDickkopfreceptorsthatregulateWnt/beta-cateninsignalling.
Nature2002;417:664-667121DavidsonG,MaoB,delBarcoBarrantesI,NiehrsC.
KremenproteinsinteractwithDickkopf1toregulateanteroposteriorCNSpatterning.
Development2002;129:5587-5596122EllwangerK,SaitoH,Clément-LacroixP,MaltryN,Nieder-meyerJ,LeeWK,BaronR,RawadiG,WestphalH,NiehrsC.
TargeteddisruptionoftheWntregulatorKremeninduceslimbdefectsandhighbonedensity.
MolCellBiol2008;28:4875-4882123WangK,ZhangY,LiX,ChenL,WangH,WuJ,ZhengJ,WuD.
CharacterizationoftheKremen-bindingsiteonDkk1andelucidationoftheroleofKremeninDkk-mediatedWntan-tagonism.
JBiolChem2008;283:23371-23375124González-SanchoJM,AguileraO,GarcíaJM,Pendás-FrancoN,PeaC,CalS,GarcíadeHerrerosA,BonillaF,MuozA.
TheWntantagonistDICKKOPF-1geneisadownstreamtargetofbeta-catenin/TCFandisdownregulatedinhumancoloncancer.
Oncogene2005;24:1098-1103125Pendás-FrancoN,GarcíaJM,PeaC,ValleN,PálmerHG,HeinniemiM,CarlbergC,JiménezB,BonillaF,MuozA,González-SanchoJM.
DICKKOPF-4isinducedbyTCF/beta-cateninandupregulatedinhumancoloncancer,promotestumourcellinvasionandangiogenesisandisrepressedby1alpha,25-dihydroxyvitaminD3.
Oncogene2008;27:4467-4477126YuB,YangX,XuY,YaoG,ShuH,LinB,HoodL,WangH,YangS,GuJ,FanJ,QinW.
ElevatedexpressionofDKK1isassociatedwithcytoplasmic/nuclearbeta-cateninaccumula-tionandpoorprognosisinhepatocellularcarcinomas.
JHepa-tol2009;50:948-957127MaoB,NiehrsC.
Kremen2modulatesDickkopf2activityduringWnt/LRP6signaling.
Gene2003;302:179-183128ChenL,WangK,ShaoY,HuangJ,LiX,ShanJ,WuD,ZhengJJ.
StructuralinsightintothemechanismsofWntsignalingantagonismbyDkk.
JBiolChem2008;283:23364-23370129HoangBH,KuboT,HealeyJH,YangR,NathanSS,KolbEA,MazzaB,MeyersPA,GorlickR.
Dickkopf3inhibitsinvasionandmotilityofSaos-2osteosarcomacellsbymodulatingtheWnt-beta-cateninpathway.
CancerRes2004;64:2734-2739130LeeEJ,JoM,RhoSB,ParkK,YooYN,ParkJ,ChaeM,ZhangW,LeeJH.
Dkk3,downregulatedincervicalcancer,functionsasanegativeregulatorofbeta-catenin.
IntJCancer2009;124:287-297131YueW,SunQ,DacicS,LandreneauRJ,SiegfriedJM,YuJ,ZhangL.
DownregulationofDkk3activatesbeta-catenin/TCF-4signalinginlungcancer.
Carcinogenesis2008;29:84-92132PatilMA,ChuaMS,PanKH,LinR,LihCJ,CheungST,HoC,LiR,FanST,CohenSN,ChenX,SoS.
Anintegrateddataanalysisapproachtocharacterizegeneshighlyexpressedinhepatocellularcarcinoma.
Oncogene2005;24:3737-3747133KwackMH,HwangSY,JangIS,ImSU,KimJO,KimMK,KimJC,SungYK.
AnalysisofcellularchangesresultingfromforcedexpressionofDickkopf-1inhepatocellularcarcinomacells.
CancerResTreat2007;39:30-36134NousoK,KobayashiY,NakamuraS,KobayashiS,TakayamaH,ToshimoriJ,KuwakiK,HagiharaH,OnishiH,MiyakeY,IkedaF,ShirahaH,TakakiA,IwasakiY,KobashiH,YamamotoK.
Prognosticimportanceoffucosylatedalpha-fetoproteininhepatocellularcarcinomapatientswithlowalpha-fetoprotein.
JGastroenterolHepatol2011;26:1195-1200135KoikeY,ShiratoriY,SatoS,ObiS,TerataniT,ImamuraM,YoshidaH,ShiinaS,OmataM.
Des-gamma-carboxypro-thrombinasausefulpredisposingfactorforthedevelopmentofportalvenousinvasioninpatientswithhepatocellularcar-cinoma:aprospectiveanalysisof227patients.
Cancer2001;91:561-569136MakinoT,YamasakiM,TakemasaI,TakenoA,NakamuraY,MiyataH,TakiguchiS,FujiwaraY,MatsuuraN,MoriM,DokiY.
Dickkopf-1expressionasamarkerforpredictingclinicaloutcomeinesophagealsquamouscellcarcinoma.
AnnSurgOncol2009;16:2058-2064137YamabukiT,TakanoA,HayamaS,IshikawaN,KatoT,MiyamotoM,ItoT,ItoH,MiyagiY,NakayamaH,FujitaM,HosokawaM,TsuchiyaE,KohnoN,KondoS,NakamuraY,DaigoY.
Dikkopf-1asanovelserologicandprognosticbio-markerforlungandesophagealcarcinomas.
CancerRes2007;67:2517-2525138SatoN,YamabukiT,TakanoA,KoinumaJ,AragakiM,Ma-sudaK,IshikawaN,KohnoN,ItoH,MiyamotoM,Nakaya-maH,MiyagiY,TsuchiyaE,KondoS,NakamuraY,DaigoY.
WntinhibitorDickkopf-1asatargetforpassivecancerim-munotherapy.
CancerRes2010;70:5326-5336139KaiserM,MiethM,LiebischP,OberlnderR,RademacherJ,JakobC,KleebergL,FleissnerC,BraendleE,PetersM,StoverD,SezerO,HeiderU.
SerumconcentrationsofDKK-1corre-latewiththeextentofbonediseaseinpatientswithmultiplemyeloma.
EurJHaematol2008;80:490-494140HeiderU,KaiserM,MiethM,LamottkeB,RademacherJ,JakobC,BraendleE,StoverD,SezerO.
SerumconcentrationsofDKK-1decreaseinpatientswithmultiplemyelomare-spondingtoanti-myelomatreatment.
EurJHaematol2009;82:31-38141FulcinitiM,TassoneP,HideshimaT,ValletS,NanjappaP,EttenbergSA,ShenZ,PatelN,TaiYT,ChauhanD,MitsiadesC,PrabhalaR,RajeN,AndersonKC,StoverDR,MunshiNC.
Anti-DKK1mAb(BHQ880)asapotentialtherapeuticagentformultiplemyeloma.
Blood2009;114:371-379142KatyalS,OliverJH,PetersonMS,FerrisJV,CarrBS,BaronRL.
Extrahepaticmetastasesofhepatocellularcarcinoma.
Ra-diology2000;216:698-703143YangB,DuZ,GaoYT,LouC,ZhangSG,BaiT,WangYJ,SongWQ.
MethylationofDickkopf-3asaprognosticfactorincirrhosis-relatedhepatocellularcarcinoma.
WorldJGastro-enterol2010;16:755-763144SatoH,SuzukiH,ToyotaM,NojimaM,MaruyamaR,SasakiS,TakagiH,SogabeY,SasakiY,IdogawaM,SonodaT,MoriM,ImaiK,TokinoT,ShinomuraY.
Frequentepigeneticinac-tivationofDICKKOPFfamilygenesinhumangastrointesti-naltumors.
Carcinogenesis2007;28:2459-2466145MaehataT,TaniguchiH,YamamotoH,NoshoK,AdachiY,MiyamotoN,MiyamotoC,AkutsuN,YamaokaS,ItohF.
TranscriptionalsilencingofDickkopfgenefamilybyCpGislandhypermethylationinhumangastrointestinalcancer.
WorldJGastroenterol2008;14:2702-2714146HirataH,HinodaY,NakajimaK,KawamotoK,KikunoN,KawakamiK,YamamuraS,UenoK,MajidS,SainiS,IshiiN,DahiyaR.
WntantagonistgeneDKK2isepigeneti-callysilencedandinhibitsrenalcancerprogressionthroughapoptoticandcellcyclepathways.
ClinCancerRes2009;15:5678-5687147DingZ,QianYB,ZhuLX,XiongQR.
PromotermethylationandmRNAexpressionofDKK-3andWIF-1inhepatocellularcarcinoma.
WorldJGastroenterol2009;15:2595-2601148JiangT,HuangL,WangS,ZhangS.
ClinicalsignificanceofserumDkk-3inpatientswithgynecologicalcancer.
JObstetGynaecolRes2010;36:769-773149UrakamiS,ShiinaH,EnokidaH,HirataH,KawamotoK,KawakamiT,KikunoN,TanakaY,MajidS,NakagawaM,IgawaM,DahiyaR.
Wntantagonistfamilygenesasbiomark-ersfordiagnosis,staging,andprognosisofrenalcellcarci-nomausingtumorandserumDNA.
ClinCancerRes2006;12:FatimaSetal.
DKKsandWnt/β-cateninsignalling325August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
com6989-6997150PineauP,NagaiH,PrigentS,WeiY,GyapayG,WeissenbachJ,TiollaisP,BuendiaMA,DejeanA.
Identificationofthreedistinctregionsofallelicdeletionsontheshortarmofchro-mosome8inhepatocellularcarcinoma.
Oncogene1999;18:3127-3134151HirataH,HinodaY,MajidS,ChenY,ZamanMS,UenoK,NakajimaK,TabatabaiZL,IshiiN,DahiyaR.
DICKKOPF-4activatesthenoncanonicalc-Jun-NH2kinasesignalingpath-waywhileinhibitingtheWnt-canonicalpathwayinhumanrenalcellcarcinoma.
Cancer2011;117:1649-1660152AustinatM,DunschR,WittekindC,TannapfelA,GebhardtR,GaunitzF.
Correlationbetweenbeta-cateninmutationsandexpressionofWnt-signalingtargetgenesinhepatocellu-larcarcinoma.
MolCancer2008;7:21153LepourceletM,ChenYN,FranceDS,WangH,CrewsP,PetersenF,BruseoC,WoodAW,ShivdasaniRA.
Small-mol-eculeantagonistsoftheoncogenicTcf/beta-cateninproteincomplex.
CancerCell2004;5:91-102154EmamiKH,NguyenC,MaH,KimDH,JeongKW,EguchiM,MoonRT,TeoJL,KimHY,MoonSH,HaJR,KahnM.
Asmallmoleculeinhibitorofbeta-catenin/CREB-bindingproteintranscription[corrected].
ProcNatlAcadSciUSA2004;101:12682-12687155ThorneCA,HansonAJ,SchneiderJ,TahinciE,OrtonD,CselenyiCS,JerniganKK,MeyersKC,HangBI,WatersonAG,KimK,MelanconB,GhiduVP,SulikowskiGA,LaFleurB,SalicA,LeeLA,MillerDM,LeeE.
Small-moleculeinhibi-tionofWntsignalingthroughactivationofcaseinkinase1α.
NatChemBiol2010;6:829-836156HeB,YouL,UematsuK,XuZ,LeeAY,MatsangouM,Mc-CormickF,JablonsDM.
AmonoclonalantibodyagainstWnt-1inducesapoptosisinhumancancercells.
Neoplasia2004;6:7-14157YouL,HeB,XuZ,UematsuK,MazieresJ,FujiiN,MikamiI,ReguartN,McIntoshJK,Kashani-SabetM,McCormickF,JablonsDM.
Ananti-Wnt-2monoclonalantibodyinducesapoptosisinmalignantmelanomacellsandinhibitstumorgrowth.
CancerRes2004;64:5385-5389158BehariJ,ZengG,OtrubaW,ThompsonMD,MullerP,Mic-senyiA,SekhonSS,LeoniL,MongaSP.
R-Etodolacdecreasesbeta-cateninlevelsalongwithsurvivalandproliferationofhepatomacells.
JHepatol2007;46:849-857159PrangeW,BreuhahnK,FischerF,ZilkensC,PietschT,Pet-meckyK,EilersR,DienesHP,SchirmacherP.
Beta-cateninaccumulationintheprogressionofhumanhepatocarcino-genesiscorrelateswithlossofE-cadherinandaccumulationofp53,butnotwithexpressionofconventionalWNT-1targetgenes.
JPathol2003;201:250-259160OvejeroC,CavardC,PérianinA,HakvoortT,VermeulenJ,GodardC,FabreM,ChafeyP,SuzukiK,RomagnoloB,YamagoeS,PerretC.
Identificationoftheleukocytecell-derivedchemotaxin2asadirecttargetgeneofbeta-cateninintheliver.
Hepatology2004;40:167-176161CavardC,TerrisB,GrimberG,ChristaL,AudardV,Rad-enen-BussiereB,SimonMT,RenardCA,BuendiaMA,PerretC.
Overexpressionofregeneratingislet-derived1alphaand3alphagenesinhumanprimarylivertumorswithbeta-cateninmutations.
Oncogene2006;25:599-608162MukhopadhyayM,GorivodskyM,ShtromS,GrinbergA,NiehrsC,MorassoMI,WestphalH.
Dkk2playsanessentialroleinthecornealfateoftheocularsurfaceepithelium.
Devel-opment2006;133:2149-2154163LiX,LiuP,LiuW,MayeP,ZhangJ,ZhangY,HurleyM,GuoC,BoskeyA,SunL,HarrisSE,RoweDW,KeHZ,WuD.
Dkk2hasaroleinterminalosteoblastdifferentiationandmineralizedmatrixformation.
NatGenet2005;37:945-952164BarrantesIdelB,Montero-PedrazuelaA,Guadao-FerrazA,ObregonMJ,MartinezdeMenaR,Gailus-DurnerV,FuchsH,FranzTJ,KalaydjievS,KlemptM,HlterS,RathkolbB,Re-inhardC,MorrealedeEscobarG,BernalJ,BuschDH,WurstW,WolfE,SchulzH,ShtromS,GreinerE,HrabédeAngelisM,WestphalH,NiehrsC.
Generationandcharacterizationofdickkopf3mutantmice.
MolCellBiol2006;26:2317-2326S-EditorTianLL-EditorWebsterJRE-EditorZhengXMFatimaSetal.
DKKsandWnt/β-cateninsignalling
Correspondenceto:JohnMLuk,Professor,CancerScienceInstituteofSingapore,NationalUniversityofSingapore,Singa-pore117456,Singapore.
jmluk@nus.
edu.
sgTelephone:+65-65164516Fax:+65-68737690Received:June13,2011Revised:July7,2011Accepted:July14,2011Publishedonline:August10,2011AbstractLivercanceristhefifthandseventhmostcommoncauseofcancerinmenandwomen,respectively.
Wnt/β-cateninsignallinghasemergedasacriticalplayerinboththedevelopmentofnormalliveraswellasanoncogenicdriverinhepatocellularcarcinoma(HCC).
Basedonthecurrentunderstanding,thisarticlesummarizesthepossiblemechanismsfortheaberrantactivationofthispathwaywithspecificfocusonHCC.
Furthermore,wewilldiscusstheroleofdickkopfs(DKKs)inregulatingWnt/β-cateninsignalling,whichispoorlyunderstoodandunderstudied.
DKKsareafamilyofsecretedpro-teinsthatcompriseatleastfourmembers,namelyDKK1-DKK4,whichactasinhibitorsofWnt/β-cateninsignalling.
Nevertheless,notallmembersantagonizeWnt/β-cateninsignalling.
Theirfunctionalsignificanceinhepatocarcinogenesisremainstobefurthercharacter-izedforwhichthesestudiesshouldprovidenewinsightsintotheregulatoryroleofDKKsinWnt/β-cateninsignal-linginhepaticcarcinogenesis.
Becauseoftheimportantoncogenicroles,thereareanincreasingnumberoftherapeuticmoleculestargetingβ-cateninandtheWnt/β-cateninpathwayforpotentialtherapyofHCC.
2011Baishideng.
Allrightsreserved.
Keywords:Dickkopf;Hepatocellularcarcinoma;Tu-mourigenesis;Wnt/β-cateninsignallingPeerreviewers:StevenARosenzweig,PhD,Professor,Depart-mentofCellandMolecularPharmacologyandExperimentalTherapeutics,MedicalUniversityofSouthCarolina,173AshleyAvenue,Charleston,SC29425,UnitedStates;EijiMiyoshi,MD,PhD,Professor,DepartmentofMolecularBiochemistryandClinicalInvestigation,ProfessorandChairman,OsakaUni-versityGraduateSchoolofMedicine,565-08711-7Yamada-okaSuita,JapanFatimaS,LeeNP,LukJM.
DickkopfsandWnt/β-cateninsig-nallinginlivercancer.
WorldJClinOncol2011;2(8):311-325Availablefrom:URL:http://www.
wjgnet.
com/2218-4333/full/v2/i8/311.
htmDOI:http://dx.
doi.
org/10.
5306/wjco.
v2.
i8.
311hepatocellularcarcinomaANDTHEUNMETMEDICALNEEDSLivercancerranksthefifthmostcommoncancerinmenandthesecondleadingcauseofcancer-relateddeath.
Inwomen,itistheseventhmostfrequentcancerandsixthleadingcauseofcancerdeath[1].
Hepatocellularcarci-noma(HCC)isthemostcommonprimarymalignancyofliver.
MenarethreetimesmorelikelytodevelopHCCthanwomenandtheincidenceincreaseswithage[2].
HCCisprevalentinAsiaandAfrica,butrecentlyitisontheriseintheWesternworldduetoanincreaseinhepatitisCvirus(HCV)infection[3].
RiskfactorsforHCCincludechronichepatitisBvirus(HBV)andHCVinfections,cirrhosis,chronicalcoholabuse,aflatoxiningestion,non-alcoholicsteatohepatitisandothermetabolicliverdiseases[4,5].
MuchofHCCoccursinthebackgroundofcirrhosis.
About80%-90%ofpatientswithcirrhosisgoontodevelopHCCeventuallyandtheremaining10%-20%ofcasesdevelopHCCwithoutcirrhosis.
Fur-thermore,HBVandHCVinfectionsincreasetheriskofdevelopingcirrhosisandlaterHCC.
AmongtheHCCREVIEWWorldJClinOncol2011August10;2(8):311-325ISSN2218-4333(online)2011Baishideng.
Allrightsreserved.
OnlineSubmissions:http://www.
wjgnet.
com/2218-4333officewjco@wjgnet.
comdoi:10.
5306/wjco.
v2.
i8.
311WorldJournalofClinicalOncologyWJCO311August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comFatimaSetal.
DKKsandWnt/β-cateninsignallingcaseswithcirrhosis,HCVinfectionhasbeenidentifiedin27%-73%andHBVinfectionin12%-55%[6,7].
HCCsuffersfromahighmortalityrateduetolackofeffectivediagnosticmethodsforearlydetectionaswellaslackoftreatmentoptionsespeciallyforthosewithad-vanceddiseaseconditions.
DespitevigorousattemptstoscreenforearlyHCCbycommonsurveillancetechniquesusingserumα-fetoprotein(AFP)andultrasoundexami-nation,earlyHCCisasymptomaticandmostHCCcasesarepresentedlatewhensurgicaltreatmentsarenotame-nable[8].
Althoughsurgicalresectionremainsthetreatmentofchoiceforpatientswithwell-preservedliverfunction,itisassociatedwithahighriskofpost-operativecompli-cationsandtumourrecurrence[7].
LivertransplantationisanothertreatmentoptionforearlyHCCbutthisislimitedbytheshortageofsuitablelivergrafts[9].
OthersurgicaltreatmentsforHCCincluderadio-frequencyablation(RFA),microwaveablation(MWA)andtrans-catheterarterialchemoembolization(TACE).
RFAandMWAtechniquesutilisehighfrequencyradio-wavesandmicro-waves,respectively,tokilltumourtissuesbyheat.
Althoughseveralstudieshavereportedbetterdisease-freesurvivalandalowerfrequencyofrecurrenceaftersurgicalresectioncomparedtoRFA[10,11],othersreportbetteroverallsurvivalanddisease-freesurvivalforHCCpatientswithmulti-nodulartumoursfollowingRFA[12].
Recently,Simoetal[13]reportednodifferenceregardingtheefficaciesofMWAandRFAprocedures.
TACEisroutinelyperformedonHCCpatientswhoarenoteli-gibleforsurgicalresectionortumourablationtechniques.
However,thesurvivalbenefitsofTACEdependoncare-fulpatientselection.
Patientswithmulti-nodularHCC,withoutvascularinvasionandnoextrahepaticmetastasesshowabetter2-yearsurvival(63%)afterTACEthanHCCpatientswithvascularinvasionundergoingTACE(31%)[14,15].
However,tumourrecurrenceisanimportantlimitationtoanyoftheHCCtreatments,andthus,un-derstandingthemolecularbiologyofHCCiscrucialforthedevelopmentofnoveltherapies.
Inrecentyears,studieshaveshedlightontheclinicalimplicationsofsignallingpathwaysinHCC,includingtheRas/Raf/MEK/ERKpathway[16],thePI3K/Akt/mTORpathway[17],theJNKpathway[18]andtheNF-κBpath-way[19].
Apromisingapproachwouldbetoidentifymo-lecularpathwaysresponsibleforinitiatingandsustainingHCCastargetsforHCCtherapy.
ThecanonicalWnt/β-cateninsignallingpathwayisanothersuchoncogenicpathway,whichisfrequentlyactivatedinHCCandisreportedtoplayapivotalroleintumourigenesis[20].
ThisarticlereviewsthecanonicalWnt/β-cateninsignallingpathwayanditsinvolvementinHCCdevelopment.
Inad-dition,antagonistsofthispathwayandtheirimplicationsinHCCaredescribedanddiscussed.
Fromthispriorknowledge,wehopetoidentifymembersofthispathwaythatcouldserveaspotentialtargetsforHCCtherapy.
OVERVIEWOFWNT/β-CATENINPATHWAYTheWnt/β-cateninpathwayisawell-conservedpath-waythatisimportantinembryonicdevelopment,cellproliferation,survival,regenerationandself-renewal[21-23].
Likewise,agreatdealofunderstandinghasbeenachievedbystudyingpathologicalspecimensandusingmousemodelsofliverdiseasestounderstandtheaberrationsofthispathwayinliverdiseasesrangingfromhepatitistoHCC[24-26].
Basedontheseearlierstudies,theWnt/β-cateninpathwayisacentralplayerinmaintainingliverhealthandisdysregulatedinhepaticcancers,whichmakesitanattractivecandidateforpotentialtherapiesofHCC.
Inanunstimulatedcell,endogenousβ-cateninisfoundattheadherensjunctions,whereitinteractswithcompo-nentsofthecadherin-associatedproteincomplexestoconfercell-celladhesionfunctions[27,28].
Ontheotherhand,surplusβ-catenininthecytoplasmisdegradedbytheac-tionofadestructioncomplexwhichconsistsofglycogensynthasekinase3β(GSK3β),Axin,adenomatouspolypo-siscoli(APC)andcaseinkinaseIα(CKIα)[29].
β-Cateninisfirstphosphorylatedatserine-45(Ser45)byCKIαtofurtherprimeitforphosphorylationbyGSK3βatSer33,Ser37andthreonine-41(Thr41).
Thephosphorylatedβ-cateninisthenubiquitinatedbyβ-transducinrepeat-containingprotein(β-TrCP)andsubsequentlydegradedbytheproteasome[30](Figure1A).
Maheretal[31]reportedthatβ-cateninphosphorylatedatSer45andnotatSer33/Ser37/Thr41ispredominantlylocatedinthenucleus,whereasβ-cateninphosphorylatedatSer33/Ser37/Thr41ismostlylocalizedtothecytoplasm.
Thisspatialsepara-tionofβ-cateninsuggeststhatphosphorylationatSer45andatSer33/Ser37/Thr41isnotnecessarilycoupled.
ItmayalsoimplythatphosphorylationatSer45byCKIαservesanotherfunction,yettobedelineated,otherthanprimingβ-cateninforfurtherphosphorylationbyGSK3β.
Fordiseasedcondition,theWnt/β-cateninsignallingpathwayisactivateduponbindingofWnttooneofthemembersofthefrizzled(FZD)familyandtolow-densitylipoproteinreceptor-relatedprotein5(LRP5)orLRP6.
TheFZDrecruitsdishevelled(Dvl)totheplasmamembrane,whichinturnrecruitsAxinandGSK3βtoLRP5/LRP6[32].
TheintercellulardomainofLRP5/LRP6containsfivereiteratedPPPSPxSmotifs,whichareduallyphosphorylatedbyGSK3βandCKIα[33].
Thephosphor-ylationofLRP5/LRP6disruptstheformationofthedestructioncomplex,therebypreventingGSK3βfromphosphorylatingβ-catenin.
Therefore,β-cateninisnotdegradedandaccumulatesinthecytoplasmfromwhereittranslocatestothenucleus.
IntheabsenceofWnt,T-cellfactor(TCF)/lymphoidenhancerfactor(LEF)re-pressesgeneexpressionbyinteractingwithco-repressorGroucho,whichpromoteshistonedeacetylationandchromatinmodellinginthenucleus[34].
Nuclearaccumu-lationofβ-catenindisplacesGrouchofromTCF/LEFandrecruitsothertranscriptionalco-activators,e.
g.
CREBbindingprotein(CBP),forupregulationoftargetgenesthatareimplicatedincellproliferation,anti-apoptosis,andangiogenesis,suchascyclinD1[35](Figure1B).
Recentstudieshavesupportedreceptor-mediatedendocytosisforWntinducedsignalling[36,37].
Specifically,Wnt3awasshowntoinducecaveolin-dependentinternalizationof312August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comLRP6,whichwouldinturnrecruitAxintoLRP6phos-phorylatedbyGSK3βandCKIα,andtherebyleadtoβ-cateninaccumulation.
Thus,caveolinplaysacriticalroleinactivatingWnt/β-cateninsignalling[37].
Veryrecently,Chairoungduaetal[38]showedanovelmechanismofreducingcytoplasmicβ-cateninlevelsindependentlyofGSK3βphosphorylationorubiquitina-tionbyexportingβ-cateninoutofthecellviaexosomes.
Exosomesarevesiclesthatforminsideendosomesandthevesiclesarethensecretedwhentheendosomesfusewiththeplasmamembrane[39].
Theseexosomesareen-richedinE-cadherinandtetraspaninproteins(CD9andCD82).
Expressionofthesetetraspaninswasshowntodecreaseβ-cateninproteinlevels,butfurtherexperimentsshowedthatE-cadherinwasalsonecessaryforβ-cateninsecretioninexosomes.
ThemolecularmechanismfortheinclusionofCD9,CD82andE-cadherininexosomeswarrantsfurtherinvestigation.
Furthermore,howthesetetraspaninsinduceexosomeformationremainstobecharacterized.
Althoughmuchremainstobeinvestigated,thisimportantandnovelmechanismoffersanalterna-tiveroutefortheregulationofWnt/β-cateninactivity,furtherhighlightingthesignificanceofkeepingtheWnt/β-cateninpathwayundercheck.
ABERRANTWNT/β-CATENINSIGNALLINGINHCCRoleofaberrantβ-cateninactivationinHCCHCCisoneofthecancerswithahighrateofdys-regulationintheWnt/β-cateninpathwayandalthough40%-70%[40-43]ofHCCpatientshavetumourswithhighlevelsofβ-cateninaccumulation,thereislittleagreementontheuseofβ-catenininprognosis.
Nuclearaccumula-tionofβ-cateninisstronglyassociatedwithβ-cateninmutations[41].
Amajorityofβ-cateninmutationsinHCCaremissensemutationsoccurringatexon3.
Thisregionisresponsibleforphosphorylationandubiquitinationofβ-catenin,andtherefore,mutationinthisregionresults13August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comAxinAPCDKK1,DKK2,DKK4LRP5/6KremenWntsFRPWIFFZDCytoplasmPlasmamembraneHDPR1Prickle-1Dvlb-cateninCKIaDKK3b-TrCPGSK3bpppppppUbUbb-cateninpb-catenindegradationbyproteasomeTCF/LEFGrouchoNucleusLRP5/6APCWntCytoplasmPlasmamembraneb-cateninGSK3bpTCF/LEFNucleusFZDAxinDvlpb-cateninb-cateninb-cateninb-cateninCBPCKIaNLKc-Myc,CyclinD1,DKK1,etc.
miR-181miR-375BAFigure1Wnt/β-cateninsignallingintheabsenceandpresenceofWntstimulus.
A:Wnt/β-cateninsignallingisregulatedbyseveralantagoniststopreventtheformationoffrizzled(FZD)-Wnt-low-densitylipoproteinreceptor-relatedprotein5/6(LRP5/6)complex.
Secretedfrizzled-relatedprotein(sFRP)andWntinhibitoryfactor(WIF)binddirectlytoWnt,whereasdickkopfs(DKKs)bindtoLRP5/6.
Furthermore,humanhomologueofDapper(HDPR1)andPrickle-1inhibittheactionofdishevelled(Dvl).
IntheabsenceofWntstimulus,β-cateninisfirstprimedforphosphorylationbycaseinkinaseIα(CKIα)followedbyphosphorylationbyglycogensynthasekinase3β(GSK3β)atthreeresidues.
Thephosphorylatedβ-cateninistargetedforubiquitinationbyβ-transducinrepeat-containingprotein(β-TrCP)andissubsequentlydegradedbytheproteasome.
Inthenucleus,T-cellfactor(TCF)/lymphoidenhancerfactor(LEF)repressestranscriptionoftheWnt/β-cateninpathwaytargetgenesbyinteractingwithco-repressorGroucho;B:WntbindstoandactivatesFZDandLRP5/6receptors.
DvlisrecruitedtotheplasmamembraneandbindstoFZD.
ThisresultsintherecruitmentofAxinandGSK3βtoLRP5/6.
LRP5/6isthenphosphorylatedbyCKIαandGSK3β,resultinginaninactivationofthedestruc-tioncomplexandleadingtoβ-cateninaccumulationinthecytoplasm.
β-cateninthensubsequentlytranslocatestothenucleuswhereitbindswithTCF/LEFandotherco-activatorse.
g.
CREBbindingprotein(CBP)tomediatetranscriptionofgenesandmicroRNAsresponsibleforproliferationandgrowth.
APC:Adenomatouspolypo-siscoli;NLK:Nemo-likekinase;p:Phosphorylated;Ub:Ubiquitinated.
FatimaSetal.
DKKsandWnt/β-cateninsignallinginstableβ-cateninthatconsequentlyaccumulatesinthenucleus.
Maoetal[44]associatednuclearβ-cateninaccu-mulationtoβ-cateninmutation,non-invasiveformoftumourandgoodprognosis.
HCCtumourswithmutantnuclearβ-cateninresultedinabetter5-yearsurvivalthanHCCtumourswithwild-typenuclearβ-cateninac-cumulation.
Thisissuggestiveofthefactthatwild-typeβ-cateninaccumulationandmutantβ-cateninaccumula-tionarenotequivalent.
Furthermore,inthisstudyof37HCCtumourswithnuclearmutantβ-cateninaccumula-tion,19mutationsoccurredatthesitesofGSK3βphos-phorylation(Ser45,Ser33,Ser37andThr41),3tumourshadβ-catenindeletions,and15mutationswerereportedatothersites.
However,severalstudieshavecorrelatednuclearβ-cateninaccumulationtotumourprogressionandpoorprognosis[43,45,46].
Kondoetal[46]reportedthatβ-cateninaccumulationandβ-cateninmutationdonotoccurearlyinhepatocarcinogenesis,butcouldbeas-sociatedwithmalignantprogressionofHCC.
Similartothesefindings,Inagawaetal[45]observedpoorprognosisinHCCpatientswithnuclearβ-cateninaccumulationingradeⅢHCCtumoursandnotingradeⅠorgradeⅡHCCtumours.
Furthermore,nuclearβ-cateninac-cumulationinHCChasalsobeencorrelatedtoKi67(amarkerfortumourcellproliferation),suggestingthatβ-cateninpromotestumourprogression[43].
Thediscrepancyinβ-cateninaccumulationandHCCprog-nosiscouldbeduetothetypeofβ-cateninmutations.
Functionalstudiesontheroleofdifferentmutationsonβ-cateninstabilitymayofferinsightsintomechanismsinvolvedinβ-cateninregulation.
Otherreasonsforthediscrepancymayincludetumourhistologyandthesizeofthetumour.
Additionally,thepresenceofβ-cateninmutationsdemonstratesdifferentphenotypicalfeaturesinHCC.
Cieplyetal[47]reportedthatHCCtumourshar-bouringamissensemutationatexon3exhibitamoreaggressivephenotypeandmaydevelopHCCwithoutcirrhosiscomparedtoHCCwithnon-mutatedβ-catenin.
Thus,β-cateninmutationsmayserveasanindependentriskfactorforthedevelopmentofHCCintheabsenceofcirrhosis.
GreatertumoursizehasalsobeenreportedinHCCtumourswithβ-cateninmutationsascomparedtothosewithoutmutationinβ-catenin[48].
Somestud-ieshavecorrelatedcytoplasmicβ-catenin(non-nuclearβ-catenin)withpoorcellulardifferentiation,largetu-moursize(>5cmindiameter)andshortdisease-freesurvival[41].
Forreasonsnotyetelucidated,HCV-associat-edHCChasagreaterfrequencyofβ-cateninmutationsthantheHBV-associatedtype[42].
Severalstudiesontransgenicanimalmodelshaveshownthatoverexpressionofmutantorstableformsofβ-cateninonitsownisnotsufficienttoinducetumoursinliver[49-51].
However,deletionofAPCinmiceresultsinhepatomegaly,hepatocytehyperplasiaandrapidmortal-ity[52].
Thus,β-cateninmutationsoraccumulationmayco-operatewithothergenesorsignallingpathwaystoresultinhepatocarcinogenesis.
However,itisalsoimportanttotakeintoaccountthefunctionalrolesofAPCthatareindependentofβ-catenin,e.
g.
APCmaintainsepithelialintegrityinnon-transformedmousemammaryepithelialcells[53]anditregulatescellcycleprogressionthroughtheSphasebyinhibitingDNAreplicationviadirectinterac-tionwithDNAincoloncancercelllines[54].
NewmousemodelsarerequiredthatmimicirregularWnt/β-cateninpathwaytounderstandtheroleofthispathwayaswellasitstherapeuticimplications.
Wnt/β-cateninpathwaytargetgenesinHCCSeveralβ-catenintargetgenesinassociationwithlivercarcinogenesiswereidentifiedbytheirhighexpressioninchronicliverdiseasesandHCC.
However,theirspecificroleinhepatocarcinogenesisremainsunknown.
Fre-quentamplificationandoverexpressionofc-MycandcyclinD1inHCCisassociatedwithcytoplasmicandnuclearβ-cateninaccumulationalongwithpoorprog-nosis[55-61].
However,thereislittleconsensusonwhethertheoverexpressionofc-MycandcyclinD1isaresultofmutationsinβ-catenin.
Cadoretetal[49]didnotreportc-MycorcyclinD1inductionintheliveroftransgenicmicethatexpressamutantformofβ-catenin,althoughsuchmicedidexhibithepatomegalyandmarkedhepa-tocellularproliferation.
Incontrast,deLaCosteetal[62]reportedactivatingsomaticmutationsinβ-cateninin50%ofhepatictumoursinc-Myctransgenicmice.
InadditiontocyclinD1andc-Myc,severalothergeneshavebeenidentifiedasdownstreammoleculesoftheWnt/β-cateninpathwayinHCC(Table1).
ExpressionofthesegeneswasdiscoveredinHCCtransgenicmiceorinHCCtissuesexhibitingaccumulationofwildtypeormutatedβ-catenin.
Forexample,glutaminesynthetaseandorphanG-protein-coupledreceptorarefrequentlyoverexpressedinHCCwithmutationinβ-catenin[63,64].
Furtherstudiesarewarrantedtounderstandwhethermutatedorstableβ-cateninresultsintranscriptionofdifferenttargetgenes,andifsilencingthesetargetgenesaffectsaberrantWnt/β-cateninpathwayinanegativefeedbackmanner.
Moreimportantly,cantheexpressionofβ-catenintargetgenesinHCC,e.
g.
glutaminesyn-thetaseorcyclinD1,besufficienttoidentifyβ-cateninactivation314August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comTable1DownstreammoleculesoftheWnt/β-cateninpathwaywithoverexpressioninhepatocellularcarcinomaGeneRef.
c-Myc[159]CyclinD1[57]Dickkopf1[126]Epidermalgrowthfactorreceptor[51]Glutaminesynthetase[63]Glutamatetransporter[63]Leukocytecell-derivedchemotaxin2[160]Ornithineaminotransferase[63]OrphanG-protein-coupledreceptor[64]Regeneratingislet-derived1α[161]Regeneratingislet-derived3α[161]FatimaSetal.
DKKsandWnt/β-cateninsignallingAlterationsofWnt/β-cateninpathwaycomponentsinHCCAccumulationofβ-catenincanalsooccurintheabsenceofβ-cateninmutationorduetoaberrantexpressionofothermembersoftheWnt/β-cateninpathway[65].
Table2summarizestheaberrantexpressionofWnt/β-cateninsig-nallingcomponentsinHCC.
WntandFZDTheWntfamilyiscomposedofnineteensecretedgly-coproteins[66].
TheybindtotheextracellulardomainofFZDsandactivatetheWnt/β-cateninpathway[67].
TendifferentFZDgeneshavebeenidentifiedinmammalsandallofthemencodeseventransmembranerecep-tors[68].
Wnt1isupregulatedinHCCtissuescomparedtoadjacentnon-tumourtissuesanditsexpressionhasbeenassociatedwithtumourrecurrence[69].
Furthermore,threeotherWntgenes(Wnt3,Wnt4andWnt5A),andthreeFZDgenes(FZD3,FZD6andFZD7)arealsoup-regulatedinHCCtissuesandpreneoplasticperitumouraltissuesascomparedwithnormallivertissues,suggestingthattheiroverexpressionmaybeanearlyeventinhepa-tocarcinogenesis.
However,onlytheoverexpressionofFZD7hasbeenassociatedwithnuclearand/orcytoplas-micaccumulationofβ-catenininHCC[70-72].
GSK3β,CKIα,AxinandAPCTherearenoreportsofmutationsoraberrantexpressionofCKIαinHCC.
Afewstudieshavereportedoverex-pressionofphosphorylatedGSK3β(phospho-GSK3β,inactiveformofGSK3β)inHCCcellsandtissues,butthishasnotbeenfoundtobeassociatedwithβ-cateninaccumulation[73,74].
TherearetwoAxingenes.
Axin1isconstitutivelyexpressed,whereasAxin2(alsoknownasAxilorCon-ductin)isinducedbyWnt/β-cateninsignallingandtakespartinanegativefeedbackloop[75].
Kimetal[76]recentlyobservedreducedAxin1expressiononlyinHCCtissuesandnotincirrhoticnodules,implyingthatitsreducedex-pressionisindependentofcirrhosis.
TherearecurrentlynoreportsofAxin2expressioninHCC.
However,Axin1andAxin2mutationsarerareinHCCoccurringinonly10%and3%ofHCCcases,respectively[48,77,78].
AmajorityofthesemutationsareframeshiftornonsensemutationsandmorethanhalfofHCCcaseswithAxin1orAxin2mutationshaveβ-cateninaccumulationinthecytoplasmornucleus[77].
Interestingly,Zucman-Rossietal[79]recentlydemonstratedthatthelossofAxin1functionisnotequivalenttogain-of-functionofβ-catenin,suggestingthatAxin1couldalsobeparticipatinginotherpathwaysinHCC.
APCmutationsareveryrareinHCCandpromotermethylationhasbeenreportedtoplayanimportantroleinitsinactivation[80].
DvlandPeptidyl-prolylcis/transisomeraseDvlisapositiveregulatorofWnt/β-cateninsignallingandpreventsGSK3βfromphosphorylatingβ-catenin,leadingtoβ-cateninstabilisation[81].
ItsoverexpressionhasbeenshowntobecriticalinWnt/β-cateninsignal-lingactivationandβ-cateninaccumulationinvariouscancersincludingHCC[82].
TwoinhibitorsofDvlhavebeenidentifiedincludingPrickle-1[82]andhumanhomo-logueofDapper[83]bothofwhicharereducedinHCCandtheirreducedexpressionhassignificantassociationwithβ-cateninaccumulation.
Peptidyl-prolylcis/transisomerase(PIN1)isanotherpositiveregulatoroftheWnt/β-cateninpathwayandfunctionsbyinhibitingtheinteractionbetweenβ-cateninandAPC[84].
Itisalsoover-expressedinmorethan50%ofHCCcasesandthishasbeencorrelatedtoincreasedβ-cateninandcyclinD1ac-cumulation.
Furthermore,β-cateninmutationandPIN1overexpressionaremutuallyexclusiveeventsinHCC,suggestingthatmechanismsotherthanβ-cateninmuta-tionalsoleadtoβ-cateninstabilisationandaccumula-tion[65].
TCF/LEFThehumanTCF/LEFfamilyconsistsoffourmembers,LEF-1,TCF-1,TCF-3andTCF-4,andallmemberscon-tainaconservedhighmobilitygroupboxtobendDNAtoallowbindingoftranscriptionfactors,aβ-catenin-bindingdomaintobindβ-cateninandatranscriptionre-pressiondomaintorecruitco-repressorslikeGroucho[85].
Whenβ-catenintranslocatestothenucleus,itbindstotheβ-catenin-bindingdomainofTCF/LEFandactivatestranscriptionoftargetgenes.
TheTCF/LEFfamilyhasseveralsplicedformswithunknownfunctionalsignifi-canceanditissuggestedthattheymayactivatepreferen-tialgenes[86].
InHCC,mutationsofTCF-4arerarewith315August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comTable2AberrantexpressionofWnt/β-cateninpathwaycom-ponentsinlivercancerComponentExpressionintumourtissuesvsnon-tumour/healthytissuesIncidence(%)MethodRef.
β-cateninHigh40-70IHC[40-43]Wnt1High41PCR[69]Wnt3High39-76PCR[70,71]Wnt4High20PCR[71]Wnt5AHigh25PCR[71]FZD3High41PCR[71]FZD6High31PCR[71]FZD7High33-90PCR[70-72]phospho-GSK3βHigh52IHC[73]Axin1Low67IHC[76]DvlHigh71Westernblotting[82]Prickle-1Low55PCR[82]HDPR1Low58PCR[83]PIN1High53Westernblotting[65]TCF-4High91PCR[87]LEF-1High52IHC[88]CDH17High72IHC[90]CDH17:Cadherin-17;Dvl:Dishevelled;FZD:Frizzled;phospho-GSK3β:Phosphorylatedglycogensynthasekinase3β;HDPR1:HumanhomologueofDapper;IHC:Immunohistochemistry;LEF:Lymphoidenhancerfactor;PCR:Polymerasechainreaction;PIN1:Peptidyl-prolylcis/transisomerase;TCF:T-cellfactor.
FatimaSetal.
DKKsandWnt/β-cateninsignallingJiangetal[87]reportingmutationsin2of32HCCcases,whereasCuietal[25]didnotreportanymutationin34HCCcases.
However,overexpressionofTCF-4inHCCtissueshasbeencorrelatedtoc-Mycoverexpression,tu-mourcapsulestatusandintrahepaticmetastasis[87].
LEF-1isalsooverexpressedinHCCandassociatedwithcyclinD1overexpression,butitisnotfoundtobecorrelatedwithanyclinicopathologicalparameters[88].
Therefore,itisnotknownwhethertheaberrantexpressionofTCF-4andLEF-1isaprimaryeventinHCCoraresultofab-errantWnt/β-cateninsignallinginHCC.
ExpressionofotherTCF/LEFfamilymembersinHCChasnotbeenreported.
Cadherin-17Cadherin-17(CDH17)belongstothecadherinsuperfam-ilycomprisingtransmembraneglycoproteinsthatmediatecalcium-dependentcell-celladhesion[89].
Sinceβ-cateninplaysanessentialroleincadherin-mediatedcell-celladhe-sionaswellasinWnt/β-cateninsignalling,ourlaboratoryidentifiedCDH17toaffectWnt/β-cateninsignallinginHCC.
CDH17isoverexpressedinHCCcellsandinmorethan70%ofHCCcases[90].
Inaddition,overexpressionofanalternativesplicevariantofCDH17(lackingexon7)inmorethan50%ofHCCiscorrelatedtotumourrecur-rence,venousinfiltrationandreducedoverallsurvival[91].
Interestingly,theCDH17splicevariantisfoundtobetriggeredbytwosinglenucleotidepolymorphismsthatareidentifiedasgeneticfactorscontributingtothehighdevel-opmentofHCCinAsia[92].
Furthermore,RNAinterfer-ence(RNAi)-mediatedknockdownofCDH17inmeta-staticHCCcellshasbeenshowntoinhibittumourigenesisinvivoandreduceWnt/β-cateninsignallingbydecreasingphospho-GSK3βandcyclinD1.
Thiswasaccompaniedbyre-localisationofβ-catenintothecytoplasm[93].
TetraspaninsTetraspaninsaretransmembraneproteinsknowntoaf-fectawiderangeoffunctionsincludingcell-celladhe-sion,cellgrowthandsuppressionofmetastasis[94].
TherecentinvolvementoftetraspaninsCD9andCD82inanovelmechanismtoantagonizeWnt/β-cateninsignallingbyexosomalreleaseofβ-cateninisanexcitingavenuetoexploreinHCC.
Thisexosomalreleaseofβ-cateninmaybecompromisedincancerswithhighWnt/β-cateninsignalling.
CD9andCD82aresuppressorsofmetastasisandtheirexpressionisreducedinHCCwithportalveininvasionand/orintrahepaticmetastasis[95].
Chairoungduaetal[38]demonstratedWnt/β-cateninsignallinginhibitioninametastaticcelllinefollowingrestorationofCD82expression.
Thus,thesetetraspaninsmaysuppressmetas-tasisbyantagonizingWnt/β-cateninsignallingbytarget-ingβ-cateninforexosomalrelease.
ItwillbeimportanttoinvestigatethecorrelationbetweenCD9andCD82withβ-catenininHCC.
MicroRNAsMicroRNAs(miRNAs)aresmallnon-codingRNAsthatregulatepost-transcriptionalgeneexpression[96].
TheyareaberrantlyexpressedinHCCcomparedtotheirnon-tumourlivertissues[97-99]andcontributetolivertumou-rigenesis[100,101].
SeveralmiRNAshavebeenidentifiedtoaffecttheWnt/β-cateninpathway[102].
Usingaglobalmicroarray-basedmiRNAprofilingapproach,Jietal[103]identifiedmiRNA-181(miR-181)tobeupregulatedinHCCtumoursthatwerepositiveforepithelialcelladhe-sionmolecule(EpCAM)andAFP(EpCAM+AFP+).
SuchtumoursdemonstratedcancerstemcellpropertiesandanactivationofWnt/β-cateninsignalling.
InvitrostudiesshowedacorrelationbetweenoverexpressionofmiR-181andβ-catenininHCCcellsandfurtherdemonstratedthatmiR-181promotedthestemnessofEpCAM+AFP+HCCcellsbytargetingCDX2(caudaltypehomeoboxtranscriptionfactor2),GATA6(GATAbindingprotein6,ahepatictranscriptionalregulatorofdifferentiation)andnemo-likekinase(NLK,aninhibitorofWnt/β-cateninsignalling).
ThesefindingsprovideevidencethatmiR-181istranscriptionallyactivatedbyWnt/β-cateninsignallingandinturninhibitsitsregulators.
Inaddition,miR-375isanothermiRNAinvolvedintheWnt/β-cateninpathwayanditisdownregulatedbyβ-catenininHCC[104].
How-ever,thefunctionofmiR-375andthemechanismsbywhichitisregulatedbyβ-cateninarenotclear.
FurtherresearchisneededtoinvestigatetheinvolvementofmiR-NAsinWnt/β-cateninsignallinginHCC.
Yes-associatedproteinTheHipposignallingpathwaycontrolsorgansizebyregulatingcellproliferationandapoptosis.
Thesignallingcascadeofthispathwayultimatelyleadstothephosphor-ylationofyes-associatedprotein(YAP),adownstreameffectorofthispathway.
YAPisatranscriptionalco-ac-tivatoranditsphosphorylationcausesittoremaininthecytoplasmandpreventthetranscriptionofgenesrespon-sibleforcellproliferationandinhibitionofapoptosis[105].
Recently,afewstudieshavedescribedtheHippopathwayasanegativeregulatorofWnt/β-cateninsignalling[106,107].
Varelasetal[106]reportedphosphorylatedTaz(componentoftheHippopathway)toinhibittheactivationofDvl,therebypreventingβ-cateninstabilisationandactivation.
Heallenetal[107]recentlyshowednuclearinteractionofunphosphorylatedYAPandβ-cateninincardiaccellsofmicewithdysregulatedHipposignalling.
ThesemicehadenlargedheartsandoverexpressedWnt/β-catenintargetgenes.
Additionally,dysregulationoftheHipposignallingpathwayandinhibitionofβ-cateninresultedinrestric-tionofcardiomyocyteovergrowth.
Thisstudyofferedinsightsintothedirectinteractionbetweenthedown-streameffectorsofthesetwoimportantpathways.
InHCC,nuclearoverexpressionofYAP(unphosphorylatedYAP)hasbeenreportedin62%ofHCCandhasbeenassociatedwithshortdisease-freesurvivalandoverallsurvival[108].
Sinceβ-cateninisalsofoundtoaccumulateinthenucleusofHCCpatients[43,45],furtherstudiesarewarrantedtounderstandtheclinicalimplicationsofYAPandβ-cateninoverexpressioninHCC.
316August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comFatimaSetal.
DKKsandWnt/β-cateninsignalling317August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comWNTSIGNALLINGANTAGONISTSAsmentionedabove,therearemanyfactorsaffectingtheWnt/β-cateninpathwayinHCC.
However,theoccur-renceofthesefactorsisnotabsoluteineachclinicalcaseofHCC.
Therefore,theinvolvementofotherfactorsinmodulatingtheWnt/β-cateninpathwayisspeculated.
FactorsupstreamofGSK3β/Axin/APCproteincom-plex,likethoseoccurringattheextracellularlevel,mayalsobethoseinvolved.
Forthis,secretedWntantagonistsaredistinguishedexamples.
TheseantagonistscanbedividedbroadlyintotwogroupsbasedontheirdifferentmechanismsinpreventingtheinteractionofWntwithLRP5/LRP6[109].
GroupⅠconsistsofWise,sclerostinanddickkopfs(DKKs)thatbinddirectlytoLRP5/LRP6.
GroupⅡcomprisesWntinhibitoryfactorandsecretedfrizzled-relatedproteinthatactbydirectlybindingtoWntproteinsandpreventtheformationoftheWntreceptorcomplex.
ThisreviewwillfocusontheroleofDKKsontheWnt/β-cateninpathwayinHCC.
StructureofDKKsTheDKKfamilyconsistsoffourmembers(DKK1toDKK4).
Theyaresecretedglycoproteinsof225-350ami-noacidswithmolecularweightsbetween25and29kDaforDKK1,DKK2andDKK4,and38kDaforDKK3.
DKKscontaintwoconservedcysteine-richdomains,eachofwhichisseparatedbyalinkerregionofvariouslengths[110].
Eachcysteine-richdomaincontainstencys-teineresidues.
Theamino-terminalcysteine-richdomain(Cys-1)isuniquetoeachDKK,whereasthecarboxyl-terminalcysteine-richdomain(Cys-2)ishighlyconservedamongallmembersoftheDKKfamily.
ThepositionofeachcysteineresidueintheCys-2domaincloselyre-semblesproteinsinthecolipasefamily[111].
Becauseoftheroleofcolipasesinlipidhydrolysis[112],thepresenceofthisfeaturesuggeststheabilityofDKKstointeractwithlipidsinregulatingWnt/β-cateninsignalling[111].
AmongallDKKs,DKK3isthemostdivergentmember[113].
Itcontainsanextendedamino-terminaldomainprecedingtheCys-1regionandanextendedcarboxyl-terminaldo-mainfollowingtheCys-2domain.
AllDKKspossesssev-eralpotentialsitesforproteolyticcleavagebyfurin-typeproteases,indicatingthattheymaybesubjectedtopost-translationalmodifications[114].
Figure2AandBillustratethedifferencesbetweendifferentDKKs.
FunctionsofDKKsontheWnt/β-cateninpathwayMembersoftheDKKfamilydiffernotonlyintheirstructuresbutalsointheirmRNAexpressioninHCCandintheirabilitytomodulateWnt/β-cateninsignalling.
Table3summarizesthesedifferencesbetweenmembersoftheDKKfamily.
DKK1isthemoststudiedmemberoftheDKKfamily.
ItwasoriginallyidentifiedasaheadinducerwhenitsmRNAwasinjectedintoXenopusembryos[115].
Experi-mentsinactivatingXenopusDKK1withanti-DKK1anti-bodies[115]andinvolvingDKK1knockoutmice[116]showalackofanteriorheadstructures,highlightingitsimpor-tanceinheadformation.
DKK1inhibitsWnt-inducedstabilisationofβ-cateninandtwomodelshavebeenpro-posed.
ThefirstmodelproposesthatDKK1bindstotheextracellulardomainofLRP5/LRP6andpreventstheformationoftheFZD-Wnt-LRP5/LRP6complexinre-sponsetoWnt,therebyattenuatingWntactivity[117].
ThesecondmodelproposesthatDKK1inhibitsWntsignal-lingbyinducingclathrin-dependentinternalisationofLRP6[37].
However,contrarytotheresultsofYamamotoetal[37],Blitzeretal[118]usedmousefibroblastcellstoshowthatclathrin-dependentinternalisationofLRP6isrequiredtopropagateWnt/β-cateninsignalling,anddisturbingthisclathrin-mediatedendocytosisblocksWntactivity.
TheserecentfindingssuggestthatmechanismsofDKK1an-tagonisticactivitymayvaryindifferentcelltypes.
Figure2Domainstructureandphylogenetictreeofhumandickkopfproteins.
A:Eachdickkopf(DKK)containstwocysteine-richdomains,eachofwhichisseparatedbyalinkerregionofvariouslengths.
Theamino-terminalcysteine-richdomain(Cys-1)domainsareuniquetoeachDKK,whereasthecarboxyl-terminalcysteine-richdomain(Cys-2)domainsareconservedamongallmembersoftheDKKfamily;B:PhylogenetictreeoftheDKKproteins.
AminoacidsequenceswerealignedbytheClustalWprogram,andthephylogenetictreewasconstructedbytheneighbourjoiningmethod.
Thescalebarindicatestheestimatednumberofsub-stitutionsper50aminoacids.
SP:Signalpeptide.
DKK30.
05DKK4DKK2DKK1DKK1DKK2DKK3DKK4266aminoacids259aminoacids350aminoacids224aminoacidsScalebar50aminoacidsBALinkerregionSPSPSPSPCys-1Cys-1Cys-1Cys-1Cys-2Cys-2Cys-2Cys-2FatimaSetal.
DKKsandWnt/β-cateninsignalling318August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comInadditiontobindingtoLRP5/LRP6,DKK1alsobindstoKremen1(Krm1)andKrm2,whichbelongtoanotherclassoftransmembranereceptors[119,120].
DKK1bindstoKrm1andKrm2withhighaffinityandthisinhibitsWnt/β-cateninsignalling[121].
Despitethat,geneknockoutstudiesinmicehaveshownthatKrm1andKrm2arenotuniversallyrequiredforDKK1-associatedfunction[122].
ItwasdemonstratedthatDKK1mutantsareabletoantagonizeWntactivitywithoutbindingtoKrm1orKrm2[123].
Therefore,KrmsmaynotbeessentialforDKK1functionandfurtherstudiesareneededtounder-standtheirinvolvementinWnt/β-cateninsignalling.
AmongallDKKs,DKK4demonstratessimilaran-tagonisticactivitytowardstheWnt/β-cateninpathwayasDKK1bybindingtoLRP5/LRP6andKrms[120].
WhilefunctioningupstreamoftheWnt/β-cateninpathway,DKK1andDKK4arealsodownstreamtargetsoftheWnt/β-cateninpathway,creatinganegativefeedbacklooptoregulateWnt/β-cateninsignalling[124,125].
However,thisfeedbackmechanismisoftenabrogatedincancersincludingHCC[125,126].
LikeDKK1andDKK4,DKK2alsobindstoLRP5/LRP6andKrms.
However,DKK2mayserveasanan-tagonistoranagonisttotheWnt/β-cateninpathwaydependingoncellularcontext.
Forinstance,overexpres-sionofDKK2in293fibroblastcellsresultsinWnt/β-cateninpathwayactivation,whereasco-transfectionofDKK2andKrm2inthesamecelltypeinhibitsthispathway[127].
Toexplainthisphenomenon,Chenetal[128]identifiedYWTDβ-propellerdomainsofLRP5/LRP6asthedockingsitesforCys-2ofmouseDKK2,whilethisCys-2domainalsocontainsbindingsiteforKrm1andKrm2.
Therefore,theexpressionofKrmsservesasaswitchforthedualroleofDKK2ontheWnt/β-cateninpathway.
UnlikeotherDKKshavingaroleinWnt/β-cateninsignallingbybindingtoLRP5/LRP6andKrms,thereceptorforDKK3hasnotbeenidentifiedanditsef-fectonthispathwayremainsunclear.
EarlierstudiesinXenopussuggestedthatDKK3isnotinvolvedinWnt/β-cateninsignalling[113,115],butrecentstudieshavedemon-stratedthatDKK3isassociatedwithareductionincyto-plasmicandnuclearaccumulationofβ-catenininSaos-2osteosarcomacells,lungcancercellsandcervicalcancercells[129-131].
Interestingly,Leeetal[130]identifiedβ-TrCPasabindingpartnertoDKK3,andpossiblyDKK3mayreduceβ-cateninlevelsviaubiquitination.
Therefore,thepotentialroleofDKK3inWnt/β-cateninpathwayre-mainstobedetermined.
SincedifferentDKKsexhibitdifferenteffectsonWnt/β-cateninactivity,itwillbeimportanttostudythemechanismsbywhicheachmemberoftheDKKfamilyexertsitseffectontheWnt/β-cateninpathway.
Further-more,asreportedbyYamamotoetal[37]andBlitzeretal[118],DKKsmayhavedifferentmechanismsofactionindif-ferentcelltypes.
Additionally,sharingthesamereceptormaynotimplyexhibitingthesameeffectasinthecaseofDKK1andDKK2.
Forinstance,theybothbindtoLRP5/LRP6andKrms,butDKK1servesasanantago-nist,whereasDKK2servesasbothanagonistandanantagonisttotheWnt/β-cateninpathway.
DKKsinHCCAsmentionedabove,theWnt/β-cateninpathwayplaysacriticalroleinHCCandnotsurprisingly,Wntinhibitors,includingDKKs,areinvolved.
MoreresearchstudieshavebeendedicatedtostudyingtheroleofDKKsinHCCoverthepastfewyears.
DKK1SeveralstudieshavereportedoverexpressionofDKK1inHCCcelllinesandtissues[126,132,133],whileYuetal[126]wasthefirsttodemonstrateacorrelationbetweenDKK1overexpressionandcytoplasmic/nuclearβ-cateninaccu-mulationinHCC.
Itwasdemonstratedthroughinvitroas-saysthatDKK1failedtoinhibitTCF-mediatedtranscrip-tionalactivityinHCCcellswithcytoplasmicornuclearβ-cateninaccumulation,suggestinganabrogationofthenegativefeedbackloopofDKK1inHCC.
SurvivalanalysiscorrelatedoverexpressionofDKK1withpoorprognosisofHCCpatients,andDKK1wasidentifiedasanindependentprognosticmarkerforoverallsurvivalandCys-1:Amino-terminalcysteine-richdomain;Cys-2:Carboxyl-terminalcysteine-richdomain;HCC:Hepatocellularcarcinoma;LRP:Lipoproteinreceptor-relatedprotein;DKK:Dickkopf.
Table3DifferencesbetweentheDKKfamilymembersPropertyDKK1DKK2DKK3DKK4Ref.
VariantsUnknownUnknown2Unknown[130]LocationofCys-185-138aminoacids78-127aminoacids147-195aminoacids41-90aminoacids[110]LocationofCys-2189-263aminoacids183-256aminoacids208-284aminoacids145-218aminoacids[110]Lengthoflinkerregion(numberofaminoacids)50561254[110]ReceptorLRP5/LRP6LRP5/LRP6NotidentifiedLRP5/LRP6[113]ActivityinWnt/β-cateninsignallinginHCCAntagonistNotreportedNotreportedNotreported[126]ExpressioninHCCtissuesOverexpressedReducedReducedNotreported[126,143]PhenotypeofknockoutmiceEmbryoniclethalityViable,fertile,blindness,osteopaeniaViable,fertile,hyperactive,increasedIgM,naturalkillercellsandhaematocritlevelsNotreported[116,162-164]FatimaSetal.
DKKsandWnt/β-cateninsignalling319August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comdisease-freesurvival.
The5-yearoverallsurvivalanddis-ease-freesurvivalratesforHCCpatientsoverexpressingDKK1(43.
4%and34.
2%,respectively)weresignificantlylowerthanHCCpatientswithreducedDKK1expres-sion(59.
3%and55.
2%,respectively).
AlthoughelevatedlevelsofAFPremainthegoldstandardforscreeningHCC,thereare,however,asubgroupofpatientswhohaveHCCandnormallevelsofAFP.
WhenpatientswerestratifiedaccordingtoAFPlevels,DKK1overexpressiondemonstratedworseprognosisforAFP-normalHCCpatients,suggestingthatDKK1mayserveasaprognosticmarkerforthisgroupofpatients.
Furthermore,HCCpatientswithhighDKK1andβ-cateninexpressionalsoshowedpoorprognosis.
The5-yearoverallsurvivalanddisease-freesurvivalrateswere66.
0%and59.
8%,respec-tively,forHCCpatientswithoutDKK1andβ-cateninexpression,and46.
0%and18.
0%forHCCpatientswithhighDKK1andhighβ-cateninexpression[126].
ThisstudyhighlightstheimportantroleofDKK1inWnt/β-cateninsignallinginHCC.
OtherthantotalAFPlevels,lensculinarisagglutinin–reactiveAFP(AFP-L3)andprothrombininducedbyvitaminKabsence-Ⅱ(PIVKA-Ⅱ)havebeenreportedastumourmarkersforHCC.
HighserumAFP-L3levelshaverecentlybeenreportedasaprognosticmarkereveninHCCpatientswithlowAFPlevels[134].
Additionally,highserumlevelsofPIVKA-ⅡhavebeenassociatedwithadvancedHCCwithportalveininvasion[135].
BecauseofitshighexpressioninHCCtissuesanditssecretorynature,DKK1ishypothesizedtobepresentathighlevelsintheserumofHCCpatients.
However,therearenostudiesevaluatinghighDKK1serumlevelsonHCCprogressionorprognosisanditwouldbeim-portanttoconductsuchastudytounderstandtheclinicalsignificanceofDKK1inHCC.
Ontheotherhand,highlevelsofDKK1inpatients'serumareassociatedwithpoorprognosisinvariouscancersincludingoesophagealsquamouscellcarcinoma[136],lungcancer[137],breastcan-cer[138]andcervicalcancer[138],suggestingthattheserumlevelofDKK1mayalsoreflecttheprognosisofHCCpatients.
Inmultiplemyeloma,highDKK1serumlevelsareassociatedwithosteolyticbonelesions[139]andpatientsrespondingtoanti-myelomatreatmentshowadecreaseinDKK1serumlevels[140],suggestingtheinvolvementofDKK1inthisaspect.
Recently,Fulcinitietal[141]evaluatedtheeffectofanti-DKK1monoclonalantibody(BHQ880)inamultiplemyelomamousemodelandfoundthatitinducedboneformationandinhibitedtumour-inducedosteolyticbonelesions.
Likemultiplemyeloma,HCCisalsoosteolyticinnaturewith20%ofHCCpatientshav-ingbonemetastasis[142],makingitimportanttoassesstheroleofDKK1inbonemetastasisinHCC.
DKK2InHCC,ahigherlevelofDKK2methylationwasde-tectedinHCCtissuescomparedtocorrespondingnon-cancerouscirrhotictissues[143],suggestingitsroleinhepatocarcinogenesis.
EpigeneticsilencingofDKK2hasalsobeenreportedingastriccancer[144],oesophagealsquamouscellcarcinoma[145]andrenalcellcarcinoma[146].
Inrenalcellcarcinoma,nosignificantrelationshipwasfoundbetweenDKK2methylationandβ-cateninexpres-sion[146].
AlthoughtheeffectofDKK2onWnt/β-cateninsignallingdependsontheexpressionofLRP5/LRP6andKrms,DKK2expressionhasnotbeenstudiedinthecontextofthesemolecules.
Furthermore,therearecurrentlynoreportsontheeffectofDKK2onWnt/β-cateninsignallinginHCC.
DKK3TherearefewreportsontheclinicalsignificanceofDKK3inHCC[143,147].
ReductioninDKK3expressionisassociatedwithincreasedfrequencyofmethylationinHCCtissuescomparedtocorrespondingnon-cancerouscirrhotictissues,implyingthatDKK3methylationmaynotbeanearlyeventinHCC,butmayfunctionintheprogressionofHCC.
Furthermore,DKK3methylationhasbeensignificantlyassociatedwithshortprogression-freesurvivalinHCCpatients[143].
TheeffectofDKK3onWnt/β-cateninsignallinghasnotbeenreportedinHCC,butreducedDKK3expressionhasbeenshowntocorrelatewithβ-cateninaccumulationinlungcancer[131].
AlthoughtherearenoreportsofDKK3levelinHCCserum,reducedDKK3serumlevelsinovariancancerareassociatedwiththepresenceoflymphnodemetastasis,andhighDKK3serumlevelshavebeenassociatedwithlargetumoursincervicalcancer[148].
Interestingly,mul-tigenemethylationstatusofacombinationofWntan-tagonistgenes,includingDKK3andothers,intheserumhavebeenproposedasmarkersfordiagnosis,stagingandprognosisofrenalcellcarcinoma[149].
AlthoughdifferentWntantagonistsmayfunctiondifferently,theircumula-tivesilencingmayserveasamolecularmarkerforcancerdetection.
DKK4DKK4isleaststudiedincancer.
Itislocatedonchromo-some8p11.
2-p11.
1andthischromosomalregionexperi-encesfrequentlossofheterozygosityinHCC[150].
ThismayexplainreducedexpressionofDKK4inHCCcelllineswithoutdetectionofDKK4methylation[144].
Cur-rentlytherearenoreportsonDKK4expressioninHCCtissuesoritseffectonWnt/β-cateninsignallinginHCC.
Recently,Hirataetal[151]reportedareductionofWnttargetgenesafterectopicexpressionofDKK4inrenalcarcinoma-derivedCakicells.
However,overexpressionofDKK4alsoresultedinactivationoftheJNKpathwayandenhancedtumourgrowthinvivo.
ThissuggeststhatDKK4maybeinvolvedinpathwaysotherthanWnt/β-cateninsignalling.
POTENTIALTHERAPEUTICTARGETSFORWNT/β-CATENINSIGNALLINGINHCCMountingevidencesuggeststheroleofβ-cateninstabi-FatimaSetal.
DKKsandWnt/β-cateninsignalling320August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comlisationinpromotingtumourproliferationinHCCpa-tients[43,152].
SuchobservationsmakeWnt/β-cateninsig-nallinganattractivetargetforcancertherapy.
Inlinewiththishypothesis,smallmoleculeinhibitorshavebeende-velopedtohinderWnt/β-cateninsignallingbydisruptingprotein-proteininteractionsofcomponentsofthispath-way.
Theseincludethefungalderivatives,PKF115-584andCGP049090,thatfunctiontoimpedetheinteractionbetweenβ-cateninandtheTCFcomplex[153].
Emamietal[154]reportedanothermoleculecalledICG-001,whichdisruptstheinteractionbetweenβ-cateninandCBP.
Morerecently,pyrviniumpamoate,ananthelminticdrug,wasshowntoinhibittheWnt/β-cateninpathwaybyallo-stericactivationofCKIαandsubsequentlyincreasedlev-elsofβ-catenindestructioncomplexsecondarytoAxinstabilisation[155].
TherapeuticantibodiesagainstWnt1andWnt2havealsodemonstratedWnt/β-cateninsignallinginhibitionandsuppressionoftumourgrowthinvivo[156,157].
β-cateninsuppressionthroughchemoprophylaxismayofferanotheralternativetherapy.
R-Etodolac(enantiomerofthenon-steroidalanti-inflammatorydrugEtodolac)reducedproliferationandsurvivaloftwoHCCcelllines(HepG2andHep3B)bydecreasingthetotalandactiveforms(dephosphorylatedatSer33/Ser37/Thr41)ofβ-catenin[158].
FurtherstudieswillbeimportanttoaccessclinicalimplicationsofthesepotentialtargetsinHCC.
PERSPECTIVESAberrantactivationofWnt/β-cateninsignallingisanimportanteventinHCCthatleadstotranscriptionofvarioustargetgenesinvolvedincarcinogenesis.
ItwillbeclinicallyrelevanttotargettheWnt/β-cateninpathwayandregulateitsactivity.
However,sincethepathwaycanbeaberrantlyactivatedbythedysfunctionofseveralgenes,itisimportanttoensurethatwhenonememberistargetedtheotherfunctionsofthesametarget,ifany,arenotdisrupted.
Forinstance,β-cateninisinvolvedintheWnt/β-cateninpathwayandplaysacriticalroleincell-celladhesion.
Therefore,careshouldbetakenindesigningreagentsfordisruptionofmoleculeswithdualfunctions.
Fortumourswithmutatedβ-catenin,oneapproachcouldbetoutilizeRNAitargetingmutantβ-cateninfortherapy,asthiswilltargetselectivelyfortumourcells,butnothealthycells.
However,mutantβ-catenin-negativetumourswithaberrantWnt/β-cateninsignallingofferthegreatestchallengeastherecouldbeamultitudeofreasonsforaberrantWnt/β-cateninactiva-tion.
Therefore,alteringWnt/β-cateninsignallingmaybeadvantageousfortherapeuticmeansbutthereareca-veatsandlimitationsinvolved.
Inlinewiththeseinvestigations,DKKshavebeendiscoveredoverthepast12yearsfortheirabilitiestoin-terfereintheWnt/β-cateninpathway.
Sincethen,muchefforthasbeendedicatedtoexploringtheirfunctionsandimplicationsinliverdevelopmentanddisease.
Dif-ferentDKKsexhibitdifferentexpressionsandfunctionsinvariouscancers.
Someimportantquestionsremaintobeanswered.
Forexample,whereareDKKslocalisedinHCCcellsIstheirexpressionassociatedwithcytoplas-mic/nuclearβ-cateninaccumulationinHCCcellsDoDKKshaveprognosticvalueinHCCpatientsDotheyexerttheireffectsonbothwildtypeandmutantβ-catenininHCCcellsDoDKKsdegradeβ-cateninviaaubiq-uitination-dependentmechanismAredifferentDKKsinvolvedindifferentstagesofHCCdevelopmentWhatisthereceptorforDKK3AreDKKsredundantintheirfunctionsFurthermore,thenegativefeedbackmecha-nismofDKK1alsowarrantsfurtherresearch.
AlthoughDKK1inhibitsWnt/β-cateninsignalling,itsoverexpres-sionhasnoeffectonHCCcellswithcytoplasmic/nuclearβ-cateninaccumulation[126].
ThismaybesuggestiveofanabrogatednegativefeedbacklooporitmayalsoimplythattheinhibitoryeffectofDKK1isonlyfunctionalun-tilreachingapointofsaturationbeyondwhichitcannotexertitsinhibitoryeffect.
Insummary,comparedtotheoverwhelmingbodyofevidenceshowingthefunctionalroleofWnt/β-cateninsignallinginHCC,thebiologicalandphysiologicalrolesofDKKsinrelationtothispath-wayremaintobediscovered.
DKKsprovideanewav-enuetoexploreandtheyaddanotherlevelofcomplexitytothealreadycomplexWnt/β-cateninsignallinginHCC.
REFERENCES1JemalA,BrayF,CenterMM,FerlayJ,WardE,FormanD.
Globalcancerstatistics.
CACancerJClin2011;61:69-902WandsJ.
Hepatocellularcarcinomaandsex.
NEnglJMed2007;357:1974-19763AltekruseSF,McGlynnKA,ReichmanME.
Hepatocellu-larcarcinomaincidence,mortality,andsurvivaltrendsintheUnitedStatesfrom1975to2005.
JClinOncol2009;27:1485-14914AschaMS,HanounehIA,LopezR,TamimiTA,FeldsteinAF,ZeinNN.
Theincidenceandriskfactorsofhepatocel-lularcarcinomainpatientswithnonalcoholicsteatohepatitis.
Hepatology2010;51:1972-19785SeveriT,vanMalensteinH,VerslypeC,vanPeltJF.
Tumorinitiationandprogressioninhepatocellularcarcinoma:riskfactors,classification,andtherapeutictargets.
ActaPharmacolSin2010;31:1409-14206KanwalF,HoangT,KramerJR,AschSM,GoetzMB,Zeri-ngueA,RichardsonP,El-SeragHB.
IncreasingprevalenceofHCCandcirrhosisinpatientswithchronichepatitisCvirusinfection.
Gastroenterology2011;140:1182-1188.
e17NguyenVT,LawMG,DoreGJ.
HepatitisB-relatedhepato-cellularcarcinoma:epidemiologicalcharacteristicsanddis-easeburden.
JViralHepat2009;16:453-4638KudoM,HanKH,KokudoN,ChengAL,ChoiBI,FuruseJ,IzumiN,ParkJW,PoonRT,SakamotoM.
LiverCancerWorkingGroupreport.
JpnJClinOncol2010;40Suppl1:i19-i279MazzaferroV,ChunYS,PoonRT,SchwartzME,YaoFY,MarshJW,BhooriS,LeeSG.
Livertransplantationforhepa-tocellularcarcinoma.
AnnSurgOncol2008;15:1001-100710KimH,RhimH,ChoiD,LimHK,KimYS,LeeWJ,JohJW.
Recurrenceandtreatmentpatterninlong-termsurvivorswithhepatocellularcarcinoma:acomparisonbetweenra-diofrequencyablationandsurgeryasafirst-linetreatment.
WorldJSurg2010;34:1881-188611VivarelliM,GuglielmiA,RuzzenenteA,CucchettiA,Bel-lusciR,CordianoC,CavallariA.
SurgicalresectionversuspercutaneousradiofrequencyablationinthetreatmentofFatimaSetal.
DKKsandWnt/β-cateninsignallingFatimaSetal.
DKKsandWnt/β-cateninsignalling321August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comhepatocellularcarcinomaoncirrhoticliver.
AnnSurg2004;240:102-10712UenoS,SakodaM,KuboF,HiwatashiK,TatenoT,BabaY,HasegawaS,TsubouchiH.
Surgicalresectionversusra-diofrequencyablationforsmallhepatocellularcarcinomaswithintheMilancriteria.
JHepatobiliaryPancreatSurg2009;16:359-36613SimoKA,SereikaSE,NewtonKN,GerberDA.
Laparoscop-ic-assistedmicrowaveablationforhepatocellularcarcinoma:Safetyandefficacyincomparisonwithradiofrequencyabla-tion.
JSurgOncol2011;Epubaheadofprint14HuppertP.
Currentconceptsintransarterialchemoembo-lizationofhepatocellularcarcinoma.
AbdomImaging2011;Epubaheadofprint15LencioniR.
Loco-regionaltreatmentofhepatocellularcarci-noma.
Hepatology2010;52:762-77316CajaL,SanchoP,BertranE,Iglesias-SerretD,GilJ,FabregatI.
OveractivationoftheMEK/ERKpathwayinlivertumorcellsconfersresistancetoTGF-{beta}-inducedcelldeaththroughimpairingup-regulationoftheNADPHoxidaseNOX4.
CancerRes2009;69:7595-760217CalvisiDF,WangC,HoC,LaduS,LeeSA,MattuS,Deste-fanisG,DeloguS,ZimmermannA,EricssonJ,BrozzettiS,StanisciaT,ChenX,DombrowskiF,EvertM.
Increasedlipogenesis,inducedbyAKT-mTORC1-RPS6signaling,promotesdevelopmentofhumanhepatocellularcarcinoma.
Gastroenterology2011;140:1071-108318ChangQ,ZhangY,BeezholdKJ,BhatiaD,ZhaoH,ChenJ,CastranovaV,ShiX,ChenF.
SustainedJNK1activationisassociatedwithalteredhistoneH3methylationsinhumanlivercancer.
JHepatol2009;50:323-33319HeY,ZhangH,YinJ,XieJ,TanX,LiuS,ZhangQ,LiC,ZhaoJ,WangH,CaoG.
IkappaBalphagenepromoterpoly-morphismsareassociatedwithhepatocarcinogenesisinpatientsinfectedwithhepatitisBvirusgenotypeC.
Carcino-genesis2009;30:1916-192220WhittakerS,MaraisR,ZhuAX.
Theroleofsignalingpath-waysinthedevelopmentandtreatmentofhepatocellularcarcinoma.
Oncogene2010;29:4989-500521ApteU,ThompsonMD,CuiS,LiuB,CieplyB,MongaSP.
Wnt/beta-cateninsignalingmediatesovalcellresponseinrodents.
Hepatology2008;47:288-29522Nejak-BowenKN,MongaSP.
Beta-cateninsignaling,liverregenerationandhepatocellularcancer:sortingthegoodfromthebad.
SeminCancerBiol2011;21:44-5823GonzalezFJ.
Roleofbeta-cateninintheadultliver.
Hepatol-ogy2006;43:650-65324HsiehA,KimHS,LimSO,YuDY,JungG.
HepatitisBviralXproteininteractswithtumorsuppressoradenomatouspol-yposiscolitoactivateWnt/β-cateninsignaling.
CancerLett2011;300:162-17225CuiJ,ZhouX,LiuY,TangZ,RomeihM.
Wntsignalinginhepatocellularcarcinoma:analysisofmutationandexpres-sionofbeta-catenin,T-cellfactor-4andglycogensynthasekinase3-betagenes.
JGastroenterolHepatol2003;18:280-28726CalvisiDF,FactorVM,LaduS,ConnerEA,ThorgeirssonSS.
Disruptionofbeta-cateninpathwayorgenomicinstabil-itydefinetwodistinctcategoriesoflivercancerintransgenicmice.
Gastroenterology2004;126:1374-138627KemlerR.
Fromcadherinstocatenins:cytoplasmicproteininteractionsandregulationofcelladhesion.
TrendsGenet1993;9:317-32128DreesF,PokuttaS,YamadaS,NelsonWJ,WeisWI.
Alpha-cateninisamolecularswitchthatbindsE-cadherin-beta-cateninandregulatesactin-filamentassembly.
Cell2005;123:903-91529SakanakaC.
Phosphorylationandregulationofbeta-cateninbycaseinkinaseIepsilon.
JBiochem2002;132:697-70330OrfordK,CrockettC,JensenJP,WeissmanAM,ByersSW.
Serinephosphorylation-regulatedubiquitinationanddegra-dationofbeta-catenin.
JBiolChem1997;272:24735-2473831MaherMT,MoR,FlozakAS,PeledON,GottardiCJ.
Beta-cateninphosphorylatedatserine45isspatiallyuncoupledfrombeta-cateninphosphorylatedintheGSK3domain:im-plicationsforsignaling.
PLoSOne2010;5:e1018432ZengX,HuangH,TamaiK,ZhangX,HaradaY,YokotaC,AlmeidaK,WangJ,DobleB,WoodgettJ,Wynshaw-BorisA,HsiehJC,HeX.
InitiationofWntsignaling:controlofWntcoreceptorLrp6phosphorylation/activationviafrizzled,di-shevelledandaxinfunctions.
Development2008;135:367-37533ZengX,TamaiK,DobleB,LiS,HuangH,HabasR,Okamu-raH,WoodgettJ,HeX.
Adual-kinasemechanismforWntco-receptorphosphorylationandactivation.
Nature2005;438:873-87734BrantjesH,BarkerN,vanEsJ,CleversH.
TCF:LadyJusticecastingthefinalverdictontheoutcomeofWntsignalling.
BiolChem2002;383:255-26135DanielsDL,WeisWI.
Beta-catenindirectlydisplacesGroucho/TLErepressorsfromTcf/LefinWnt-mediatedtranscriptionactivation.
NatStructMolBiol2005;12:364-37136YamamotoH,KomekadoH,KikuchiA.
Caveolinisneces-saryforWnt-3a-dependentinternalizationofLRP6andac-cumulationofbeta-catenin.
DevCell2006;11:213-22337YamamotoH,SakaneH,YamamotoH,MichiueT,KikuchiA.
Wnt3aandDkk1regulatedistinctinternalizationpathwaysofLRP6totunetheactivationofbeta-cateninsignaling.
DevCell2008;15:37-4838ChairoungduaA,SmithDL,PochardP,HullM,CaplanMJ.
Exosomereleaseofβ-catenin:anovelmechanismthatan-tagonizesWntsignaling.
JCellBiol2010;190:1079-109139LotvallJ,ValadiH.
CelltocellsignallingviaexosomesthroughesRNA.
CellAdhMigr2007;1:156-15840SuzukiT,YanoH,NakashimaY,NakashimaO,KojiroM.
Beta-cateninexpressioninhepatocellularcarcinoma:apos-sibleparticipationofbeta-catenininthededifferentiationprocess.
JGastroenterolHepatol2002;17:994-100041WongCM,FanST,NgIO.
beta-Cateninmutationandover-expressioninhepatocellularcarcinoma:clinicopathologicandprognosticsignificance.
Cancer2001;92:136-14542HuangH,FujiiH,SankilaA,Mahler-AraujoBM,MatsudaM,CathomasG,OhgakiH.
Beta-cateninmutationsarefrequentinhumanhepatocellularcarcinomasassociatedwithhepati-tisCvirusinfection.
AmJPathol1999;155:1795-180143NhieuJT,RenardCA,WeiY,CherquiD,ZafraniES,Buen-diaMA.
Nuclearaccumulationofmutatedbeta-catenininhepatocellularcarcinomaisassociatedwithincreasedcellproliferation.
AmJPathol1999;155:703-71044MaoTL,ChuJS,JengYM,LaiPL,HsuHC.
Expressionofmutantnuclearbeta-catenincorrelateswithnon-invasivehepatocellularcarcinoma,absenceofportalveinspread,andgoodprognosis.
JPathol2001;193:95-10145InagawaS,ItabashiM,AdachiS,KawamotoT,HoriM,ShimazakiJ,YoshimiF,FukaoK.
Expressionandprognosticrolesofbeta-catenininhepatocellularcarcinoma:correlationwithtumorprogressionandpostoperativesurvival.
ClinCancerRes2002;8:450-45646KondoY,KanaiY,SakamotoM,GendaT,MizokamiM,UedaR,HirohashiS.
Beta-cateninaccumulationandmuta-tionofexon3ofthebeta-cateningeneinhepatocellularcar-cinoma.
JpnJCancerRes1999;90:1301-130947CieplyB,ZengG,Proverbs-SinghT,GellerDA,MongaSP.
Uniquephenotypeofhepatocellularcancerswithexon-3mutationsinbeta-cateningene.
Hepatology2009;49:821-83148Laurent-PuigP,LegoixP,BluteauO,BelghitiJ,FrancoD,Bi-notF,MongesG,ThomasG,Bioulac-SageP,Zucman-RossiJ.
Geneticalterationsassociatedwithhepatocellularcarcino-masdefinedistinctpathwaysofhepatocarcinogenesis.
Gas-troenterology2001;120:1763-177349CadoretA,OvejeroC,Saadi-KheddouciS,SouilE,FabreM,RomagnoloB,KahnA,PerretC.
Hepatomegalyintransgen-FatimaSetal.
DKKsandWnt/β-cateninsignalling322August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comicmiceexpressinganoncogenicformofbeta-catenin.
CancerRes2001;61:3245-324950HaradaN,MiyoshiH,MuraiN,OshimaH,TamaiY,Oshi-maM,TaketoMM.
Lackoftumorigenesisinthemouseliverafteradenovirus-mediatedexpressionofadominantstablemutantofbeta-catenin.
CancerRes2002;62:1971-197751TanX,ApteU,MicsenyiA,KotsagrelosE,LuoJH,Ranga-nathanS,MongaDK,BellA,MichalopoulosGK,MongaSP.
Epidermalgrowthfactorreceptor:anoveltargetoftheWnt/beta-cateninpathwayinliver.
Gastroenterology2005;129:285-30252ColnotS,DecaensT,Niwa-KawakitaM,GodardC,HamardG,KahnA,GiovanniniM,PerretC.
Liver-targeteddisrup-tionofApcinmiceactivatesbeta-cateninsignalingandleadstohepatocellularcarcinomas.
ProcNatlAcadSciUSA2004;101:17216-1722153ProsperiJR,BecherKR,WillsonTA,CollinsMH,WitteDP,GossKH.
TheAPCtumorsuppressorisrequiredforepi-thelialintegrityinthemousemammarygland.
JCellPhysiol2009;220:319-33154QianJ,SarnaikAA,BonneyTM,KeirseyJ,CombsKA,Stei-gerwaldK,AcharyaS,BehbehaniGK,BartonMC,LowyAM,GrodenJ.
TheAPCtumorsuppressorinhibitsDNAreplicationbydirectlybindingtoDNAviaitscarboxylter-minus.
Gastroenterology2008;135:152-16255KawateS,FukusatoT,OhwadaS,WatanukiA,MorishitaY.
Amplificationofc-mycinhepatocellularcarcinoma:cor-relationwithclinicopathologicfeatures,proliferativeactivityandp53overexpression.
Oncology1999;57:157-16356GotohJ,ObataM,YoshieM,KasaiS,OgawaK.
CyclinD1over-expressioncorrelateswithbeta-cateninactivation,butnotwithH-rasmutations,andphosphorylationofAkt,GSK3betaandERK1/2inmousehepaticcarcinogenesis.
Carcinogenesis2003;24:435-44257ChanKL,GuanXY,NgIO.
High-throughputtissuemicroar-rayanalysisofc-mycactivationinchronicliverdiseasesandhepatocellularcarcinoma.
HumPathol2004;35:1324-133158SatoY,ItohF,HareyamaM,SatohM,HinodaY,SetoM,UedaR,ImaiK.
AssociationofcyclinD1expressionwithfactorscorrelatedwithtumorprogressioninhumanhepato-cellularcarcinoma.
JGastroenterol1999;34:486-49359NishidaN,FukudaY,KomedaT,KitaR,SandoT,Furu-kawaM,AmenomoriM,ShibagakiI,NakaoK,IkenagaM.
AmplificationandoverexpressionofthecyclinD1geneinaggressivehumanhepatocellularcarcinoma.
CancerRes1994;54:3107-311060JooM,KangYK,KimMR,LeeHK,JangJJ.
CyclinD1over-expressioninhepatocellularcarcinoma.
Liver2001;21:89-9561UetaT,IkeguchiM,HirookaY,KaibaraN,TeradaT.
Beta-cateninandcyclinD1expressioninhumanhepatocellularcarcinoma.
OncolRep2002;9:1197-120362deLaCosteA,RomagnoloB,BilluartP,RenardCA,Buen-diaMA,SoubraneO,FabreM,ChellyJ,BeldjordC,KahnA,PerretC.
Somaticmutationsofthebeta-cateningenearefre-quentinmouseandhumanhepatocellularcarcinomas.
ProcNatlAcadSciUSA1998;95:8847-885163CadoretA,OvejeroC,TerrisB,SouilE,LévyL,LamersWH,KitajewskiJ,KahnA,PerretC.
Newtargetsofbeta-cateninsignalingintheliverareinvolvedintheglutaminemetabo-lism.
Oncogene2002;21:8293-830164YamamotoY,SakamotoM,FujiiG,TsuijiH,KenetakaK,AsakaM,HirohashiS.
OverexpressionoforphanG-protein-coupledreceptor,Gpr49,inhumanhepatocellularcarci-nomaswithbeta-cateninmutations.
Hepatology2003;37:528-53365PangR,YuenJ,YuenMF,LaiCL,LeeTK,ManK,PoonRT,FanST,WongCM,NgIO,KwongYL,TseE.
PIN1overex-pressionandbeta-cateningenemutationsaredistinctonco-geniceventsinhumanhepatocellularcarcinoma.
Oncogene2004;23:4182-418666NusseR,VarmusHE.
Wntgenes.
Cell1992;69:1073-108767MoonRT,BrownJD,TorresM.
WNTsmodulatecellfateandbehaviorduringvertebratedevelopment.
TrendsGenet1997;13:157-16268KolligsFT,BommerG,GkeB.
Wnt/beta-catenin/tcfsig-naling:acriticalpathwayingastrointestinaltumorigenesis.
Digestion2002;66:131-14469LeeHH,UenYH,TianYF,SunCS,SheuMJ,KuoHT,KoayLB,LinCY,TzengCC,ChengCJ,TangLY,TsaiSL,WangAH.
Wnt-1proteinasaprognosticbiomarkerforhepatitisB-relatedandhepatitisC-relatedhepatocellularcarcinomaaftersurgery.
CancerEpidemiolBiomarkersPrev2009;18:1562-156970KimM,LeeHC,TsedensodnomO,HartleyR,LimYS,YuE,MerleP,WandsJR.
FunctionalinteractionbetweenWnt3andFrizzled-7leadstoactivationoftheWnt/beta-cateninsignalingpathwayinhepatocellularcarcinomacells.
JHepa-tol2008;48:780-79171BengocheaA,deSouzaMM,LefranoisL,LeRouxE,GalyO,CheminI,KimM,WandsJR,TrepoC,HainautP,ScoazecJY,VitvitskiL,MerleP.
CommondysregulationofWnt/Frizzledreceptorelementsinhumanhepatocellularcarci-noma.
BrJCancer2008;99:143-15072MerleP,delaMonteS,KimM,HerrmannM,TanakaS,VonDemBusscheA,KewMC,TrepoC,WandsJR.
Functionalconsequencesoffrizzled-7receptoroverexpressioninhu-manhepatocellularcarcinoma.
Gastroenterology2004;127:1110-112273BanKC,SinghH,KrishnanR,SeowHF.
GSK-3betaphos-phorylationandalterationofbeta-catenininhepatocellularcarcinoma.
CancerLett2003;199:201-20874Desbois-MouthonC,Blivet-VanEggelpolMJ,BeurelE,BoissanM,DeléloR,CadoretA,CapeauJ.
Dysregulationofglycogensynthasekinase-3betasignalinginhepatocellularcarcinomacells.
Hepatology2002;36:1528-153675JhoEH,ZhangT,DomonC,JooCK,FreundJN,CostantiniF.
Wnt/beta-catenin/TcfsignalinginducesthetranscriptionofAxin2,anegativeregulatorofthesignalingpathway.
MolCellBiol2002;22:1172-118376KimYD,ParkCH,KimHS,ChoiSK,RewJS,KimDY,KohYS,JeungKW,LeeKH,LeeJS,JuhngSW,LeeJH.
GeneticalterationsofWntsignalingpathway-associatedgenesinhepatocellularcarcinoma.
JGastroenterolHepatol2008;23:110-11877TaniguchiK,RobertsLR,AdercaIN,DongX,QianC,MurphyLM,NagorneyDM,BurgartLJ,RochePC,SmithDI,RossJA,LiuW.
Mutationalspectrumofbeta-catenin,AXIN1,andAXIN2inhepatocellularcarcinomasandhepa-toblastomas.
Oncogene2002;21:4863-487178SatohS,DaigoY,FurukawaY,KatoT,MiwaN,NishiwakiT,KawasoeT,IshiguroH,FujitaM,TokinoT,SasakiY,ImaokaS,MurataM,ShimanoT,YamaokaY,NakamuraY.
AXIN1mutationsinhepatocellularcarcinomas,andgrowthsup-pressionincancercellsbyvirus-mediatedtransferofAXIN1.
NatGenet2000;24:245-25079Zucman-RossiJ,BenhamoucheS,GodardC,BoyaultS,GrimberG,BalabaudC,CunhaAS,Bioulac-SageP,PerretC.
DifferentialeffectsofinactivatedAxin1andactivatedbeta-cateninmutationsinhumanhepatocellularcarcinomas.
On-cogene2007;26:774-78080UmTH,KimH,OhBK,KimMS,KimKS,JungG,ParkYN.
AberrantCpGislandhypermethylationindysplasticnodulesandearlyHCCofhepatitisBvirus-relatedhumanmultistephepatocarcinogenesis.
JHepatol2011;54:939-94781KishidaS,YamamotoH,HinoS,IkedaS,KishidaM,Kiku-chiA.
DIXdomainsofDvlandaxinarenecessaryforproteininteractionsandtheirabilitytoregulatebeta-cateninstabil-ity.
MolCellBiol1999;19:4414-442282ChanDW,ChanCY,YamJW,ChingYP,NgIO.
Prickle-1negativelyregulatesWnt/beta-cateninpathwaybypromot-FatimaSetal.
DKKsandWnt/β-cateninsignalling323August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comingDishevelledubiquitination/degradationinlivercancer.
Gastroenterology2006;131:1218-122783YauTO,ChanCY,ChanKL,LeeMF,WongCM,FanST,NgIO.
HDPR1,anovelinhibitoroftheWNT/beta-cateninsig-naling,isfrequentlydownregulatedinhepatocellularcarci-noma:involvementofmethylation-mediatedgenesilencing.
Oncogene2005;24:1607-161484RyoA,NakamuraM,WulfG,LiouYC,LuKP.
Pin1regu-latesturnoverandsubcellularlocalizationofbeta-cateninbyinhibitingitsinteractionwithAPC.
NatCellBiol2001;3:793-80185WatermanML.
Lymphoidenhancerfactor/Tcellfactorex-pressionincolorectalcancer.
CancerMetastasisRev2004;23:41-5286VandeWeteringM,CastropJ,KorinekV,CleversH.
Exten-sivealternativesplicinganddualpromoterusagegenerateTcf-1proteinisoformswithdifferentialtranscriptioncontrolproperties.
MolCellBiol1996;16:745-75287JiangY,ZhouXD,LiuYK,WuX,HuangXW.
AssociationofhTcf-4geneexpressionandmutationwithclinicopathologi-calcharacteristicsofhepatocellularcarcinoma.
WorldJGas-troenterol2002;8:804-80788Schmitt-GraeffA,Ertelt-HeitzmannV,AllgaierHP,OlschewskiM,NitschkeR,HaxelmansS,KoelbleK,BehrensJ,BlumHE.
CoordinatedexpressionofcyclinD1andLEF-1/TCFtranscriptionfactorisrestrictedtoasubsetofhepatocel-lularcarcinoma.
LiverInt2005;25:839-84789LeeNP,PoonRT,ShekFH,NgIO,LukJM.
Roleofcad-herin-17inoncogenesisandpotentialtherapeuticimplica-tionsinhepatocellularcarcinoma.
BiochimBiophysActa2010;1806:138-14590WongBW,LukJM,NgIO,HuMY,LiuKD,FanST.
Identi-ficationofliver-intestinecadherininhepatocellularcarcino-ma--apotentialdiseasemarker.
BiochemBiophysResCommun2003;311:618-62491WangXQ,LukJM,LeungPP,WongBW,StanbridgeEJ,FanST.
AlternativemRNAsplicingofliverintestine-cadherininhepatocellularcarcinoma.
ClinCancerRes2005;11:483-48992WangXQ,LukJM,Garcia-BarceloM,MiaoX,LeungPP,HoDW,CheungST,LamBY,CheungCK,WongAS,LauSS,SoMT,YuWC,CaiQ,LiuKS,HuiCK,LauGK,PoonRT,WongJ,FanST.
Liverintestine-cadherin(CDH17)haplotypeisassociatedwithincreasedriskofhepatocellularcarcinoma.
ClinCancerRes2006;12:5248-525293LiuLX,LeeNP,ChanVW,XueW,ZenderL,ZhangC,MaoM,DaiH,WangXL,XuMZ,LeeTK,NgIO,ChenY,KungHF,LoweSW,PoonRT,WangJH,LukJM.
Targetingcadherin-17inactivatesWntsignalingandinhibitstumorgrowthinlivercarcinoma.
Hepatology2009;50:1453-146394ZllerM.
Tetraspanins:pushandpullinsuppressingandpromotingmetastasis.
NatRevCancer2009;9:40-5595KanetakaK,SakamotoM,YamamotoY,YamasakiS,LanzaF,KanematsuT,HirohashiS.
OverexpressionoftetraspaninCO-029inhepatocellularcarcinoma.
JHepatol2001;35:637-64296JiJ,WangXW.
Newkidsontheblock:diagnosticandprog-nosticmicroRNAsinhepatocellularcarcinoma.
CancerBiolTher2009;8:1686-169397BoutzDR,CollinsPJ,SureshU,LuM,RamírezCM,Fernán-dez-HernandoC,HuangY,AbreuRdeS,LeSY,ShapiroBA,LiuAM,LukJM,AldredSF,TrinkleinND,MarcotteEM,PenalvaLO.
Two-tieredapproachidentifiesanetworkofcancerandliverdisease-relatedgenesregulatedbymiR-122.
JBiolChem2011;286:18066-1807898BurchardJ,ZhangC,LiuAM,PoonRT,LeeNP,WongKF,ShamPC,LamBY,FergusonMD,TokiwaG,SmithR,Lee-sonB,BeardR,LambJR,LimL,MaoM,DaiH,LukJM.
mi-croRNA-122asaregulatorofmitochondrialmetabolicgenenetworkinhepatocellularcarcinoma.
MolSystBiol2010;6:40299LiuAM,ZhangC,BurchardJ,FanST,WongKF,DaiH,PoonRT,LukJM.
GlobalregulationonmicroRNAinhepati-tisBvirus-associatedhepatocellularcarcinoma.
OMICS2011;15:187-191100PineauP,VoliniaS,McJunkinK,MarchioA,BattistonC,TerrisB,MazzaferroV,LoweSW,CroceCM,DejeanA.
miR-221overexpressioncontributestolivertumorigenesis.
ProcNatlAcadSciUSA2010;107:264-269101WongCC,WongCM,TungEK,AuSL,LeeJM,PoonRT,ManK,NgIO.
ThemicroRNAmiR-139suppressesmetas-tasisandprogressionofhepatocellularcarcinomabydown-regulatingRho-kinase2.
Gastroenterology2011;140:322-331102HuangK,ZhangJX,HanL,YouYP,JiangT,PuPY,KangCS.
MicroRNArolesinbeta-cateninpathway.
MolCancer2010;9:252103JiJ,YamashitaT,BudhuA,ForguesM,JiaHL,LiC,DengC,WauthierE,ReidLM,YeQH,QinLX,YangW,WangHY,TangZY,CroceCM,WangXW.
IdentificationofmicroR-NA-181bygenome-widescreeningasacriticalplayerinEpCAM-positivehepaticcancerstemcells.
Hepatology2009;50:472-480104LadeiroY,CouchyG,BalabaudC,Bioulac-SageP,PelletierL,RebouissouS,Zucman-RossiJ.
MicroRNAprofilinginhepatocellulartumorsisassociatedwithclinicalfeaturesandoncogene/tumorsuppressorgenemutations.
Hepatology2008;47:1955-1963105LiuAM,XuMZ,ChenJ,PoonRT,LukJM.
TargetingYAPandHipposignalingpathwayinlivercancer.
ExpertOpinTherTargets2010;14:855-868106VarelasX,MillerBW,SopkoR,SongS,GregorieffA,Fel-louseFA,SakumaR,PawsonT,HunzikerW,McNeillH,WranaJL,AttisanoL.
TheHippopathwayregulatesWnt/beta-cateninsignaling.
DevCell2010;18:579-591107HeallenT,ZhangM,WangJ,Bonilla-ClaudioM,KlysikE,JohnsonRL,MartinJF.
HippopathwayinhibitsWntsignal-ingtorestraincardiomyocyteproliferationandheartsize.
Science2011;332:458-461108XuMZ,YaoTJ,LeeNP,NgIO,ChanYT,ZenderL,LoweSW,PoonRT,LukJM.
Yes-associatedproteinisanindepen-dentprognosticmarkerinhepatocellularcarcinoma.
Cancer2009;115:4576-4585109KawanoY,KyptaR.
SecretedantagonistsoftheWntsignal-lingpathway.
JCellSci2003;116:2627-2634110KrupnikVE,SharpJD,JiangC,RobisonK,ChickeringTW,AmaravadiL,BrownDE,GuyotD,MaysG,LeibyK,ChangB,DuongT,GoodearlAD,GearingDP,SokolSY,McCarthySA.
FunctionalandstructuraldiversityofthehumanDick-kopfgenefamily.
Gene1999;238:301-313111AravindL,KooninEV.
Acolipasefoldinthecarboxy-termi-naldomainoftheWntantagonists--theDickkopfs.
CurrBiol1998;8:R477-R478112vanTilbeurghH,BezzineS,CambillauC,VergerR,CarrièreF.
Colipase:structureandinteractionwithpancreaticlipase.
BiochimBiophysActa1999;1441:173-184113NiehrsC.
FunctionandbiologicalrolesoftheDickkopffam-ilyofWntmodulators.
Oncogene2006;25:7469-7481114NakayamaK.
Furin:amammaliansubtilisin/Kex2p-likeendoproteaseinvolvedinprocessingofawidevarietyofprecursorproteins.
BiochemJ1997;327(Pt3):625-635115GlinkaA,WuW,DeliusH,MonaghanAP,BlumenstockC,NiehrsC.
Dickkopf-1isamemberofanewfamilyofse-cretedproteinsandfunctionsinheadinduction.
Nature1998;391:357-362116MukhopadhyayM,ShtromS,Rodriguez-EstebanC,ChenL,TsukuiT,GomerL,DorwardDW,GlinkaA,GrinbergA,HuangSP,NiehrsC,IzpisúaBelmonteJC,WestphalH.
Dickkopf1isrequiredforembryonicheadinductionandlimbmorphogenesisinthemouse.
DevCell2001;1:423-434117SemnovMV,TamaiK,BrottBK,KühlM,SokolS,HeX.
HeadinducerDickkopf-1isaligandforWntcoreceptorFatimaSetal.
DKKsandWnt/β-cateninsignalling324August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
comLRP6.
CurrBiol2001;11:951-961118BlitzerJT,NusseR.
AcriticalroleforendocytosisinWntsig-naling.
BMCCellBiol2006;7:28119NakamuraT,NakamuraT,MatsumotoK.
ThefunctionsandpossiblesignificanceofKremenasthegatekeeperofWntsig-nallingindevelopmentandpathology.
JCellMolMed2008;12:391-408120MaoB,WuW,DavidsonG,MarholdJ,LiM,MechlerBM,DeliusH,HoppeD,StannekP,WalterC,GlinkaA,NiehrsC.
KremenproteinsareDickkopfreceptorsthatregulateWnt/beta-cateninsignalling.
Nature2002;417:664-667121DavidsonG,MaoB,delBarcoBarrantesI,NiehrsC.
KremenproteinsinteractwithDickkopf1toregulateanteroposteriorCNSpatterning.
Development2002;129:5587-5596122EllwangerK,SaitoH,Clément-LacroixP,MaltryN,Nieder-meyerJ,LeeWK,BaronR,RawadiG,WestphalH,NiehrsC.
TargeteddisruptionoftheWntregulatorKremeninduceslimbdefectsandhighbonedensity.
MolCellBiol2008;28:4875-4882123WangK,ZhangY,LiX,ChenL,WangH,WuJ,ZhengJ,WuD.
CharacterizationoftheKremen-bindingsiteonDkk1andelucidationoftheroleofKremeninDkk-mediatedWntan-tagonism.
JBiolChem2008;283:23371-23375124González-SanchoJM,AguileraO,GarcíaJM,Pendás-FrancoN,PeaC,CalS,GarcíadeHerrerosA,BonillaF,MuozA.
TheWntantagonistDICKKOPF-1geneisadownstreamtargetofbeta-catenin/TCFandisdownregulatedinhumancoloncancer.
Oncogene2005;24:1098-1103125Pendás-FrancoN,GarcíaJM,PeaC,ValleN,PálmerHG,HeinniemiM,CarlbergC,JiménezB,BonillaF,MuozA,González-SanchoJM.
DICKKOPF-4isinducedbyTCF/beta-cateninandupregulatedinhumancoloncancer,promotestumourcellinvasionandangiogenesisandisrepressedby1alpha,25-dihydroxyvitaminD3.
Oncogene2008;27:4467-4477126YuB,YangX,XuY,YaoG,ShuH,LinB,HoodL,WangH,YangS,GuJ,FanJ,QinW.
ElevatedexpressionofDKK1isassociatedwithcytoplasmic/nuclearbeta-cateninaccumula-tionandpoorprognosisinhepatocellularcarcinomas.
JHepa-tol2009;50:948-957127MaoB,NiehrsC.
Kremen2modulatesDickkopf2activityduringWnt/LRP6signaling.
Gene2003;302:179-183128ChenL,WangK,ShaoY,HuangJ,LiX,ShanJ,WuD,ZhengJJ.
StructuralinsightintothemechanismsofWntsignalingantagonismbyDkk.
JBiolChem2008;283:23364-23370129HoangBH,KuboT,HealeyJH,YangR,NathanSS,KolbEA,MazzaB,MeyersPA,GorlickR.
Dickkopf3inhibitsinvasionandmotilityofSaos-2osteosarcomacellsbymodulatingtheWnt-beta-cateninpathway.
CancerRes2004;64:2734-2739130LeeEJ,JoM,RhoSB,ParkK,YooYN,ParkJ,ChaeM,ZhangW,LeeJH.
Dkk3,downregulatedincervicalcancer,functionsasanegativeregulatorofbeta-catenin.
IntJCancer2009;124:287-297131YueW,SunQ,DacicS,LandreneauRJ,SiegfriedJM,YuJ,ZhangL.
DownregulationofDkk3activatesbeta-catenin/TCF-4signalinginlungcancer.
Carcinogenesis2008;29:84-92132PatilMA,ChuaMS,PanKH,LinR,LihCJ,CheungST,HoC,LiR,FanST,CohenSN,ChenX,SoS.
Anintegrateddataanalysisapproachtocharacterizegeneshighlyexpressedinhepatocellularcarcinoma.
Oncogene2005;24:3737-3747133KwackMH,HwangSY,JangIS,ImSU,KimJO,KimMK,KimJC,SungYK.
AnalysisofcellularchangesresultingfromforcedexpressionofDickkopf-1inhepatocellularcarcinomacells.
CancerResTreat2007;39:30-36134NousoK,KobayashiY,NakamuraS,KobayashiS,TakayamaH,ToshimoriJ,KuwakiK,HagiharaH,OnishiH,MiyakeY,IkedaF,ShirahaH,TakakiA,IwasakiY,KobashiH,YamamotoK.
Prognosticimportanceoffucosylatedalpha-fetoproteininhepatocellularcarcinomapatientswithlowalpha-fetoprotein.
JGastroenterolHepatol2011;26:1195-1200135KoikeY,ShiratoriY,SatoS,ObiS,TerataniT,ImamuraM,YoshidaH,ShiinaS,OmataM.
Des-gamma-carboxypro-thrombinasausefulpredisposingfactorforthedevelopmentofportalvenousinvasioninpatientswithhepatocellularcar-cinoma:aprospectiveanalysisof227patients.
Cancer2001;91:561-569136MakinoT,YamasakiM,TakemasaI,TakenoA,NakamuraY,MiyataH,TakiguchiS,FujiwaraY,MatsuuraN,MoriM,DokiY.
Dickkopf-1expressionasamarkerforpredictingclinicaloutcomeinesophagealsquamouscellcarcinoma.
AnnSurgOncol2009;16:2058-2064137YamabukiT,TakanoA,HayamaS,IshikawaN,KatoT,MiyamotoM,ItoT,ItoH,MiyagiY,NakayamaH,FujitaM,HosokawaM,TsuchiyaE,KohnoN,KondoS,NakamuraY,DaigoY.
Dikkopf-1asanovelserologicandprognosticbio-markerforlungandesophagealcarcinomas.
CancerRes2007;67:2517-2525138SatoN,YamabukiT,TakanoA,KoinumaJ,AragakiM,Ma-sudaK,IshikawaN,KohnoN,ItoH,MiyamotoM,Nakaya-maH,MiyagiY,TsuchiyaE,KondoS,NakamuraY,DaigoY.
WntinhibitorDickkopf-1asatargetforpassivecancerim-munotherapy.
CancerRes2010;70:5326-5336139KaiserM,MiethM,LiebischP,OberlnderR,RademacherJ,JakobC,KleebergL,FleissnerC,BraendleE,PetersM,StoverD,SezerO,HeiderU.
SerumconcentrationsofDKK-1corre-latewiththeextentofbonediseaseinpatientswithmultiplemyeloma.
EurJHaematol2008;80:490-494140HeiderU,KaiserM,MiethM,LamottkeB,RademacherJ,JakobC,BraendleE,StoverD,SezerO.
SerumconcentrationsofDKK-1decreaseinpatientswithmultiplemyelomare-spondingtoanti-myelomatreatment.
EurJHaematol2009;82:31-38141FulcinitiM,TassoneP,HideshimaT,ValletS,NanjappaP,EttenbergSA,ShenZ,PatelN,TaiYT,ChauhanD,MitsiadesC,PrabhalaR,RajeN,AndersonKC,StoverDR,MunshiNC.
Anti-DKK1mAb(BHQ880)asapotentialtherapeuticagentformultiplemyeloma.
Blood2009;114:371-379142KatyalS,OliverJH,PetersonMS,FerrisJV,CarrBS,BaronRL.
Extrahepaticmetastasesofhepatocellularcarcinoma.
Ra-diology2000;216:698-703143YangB,DuZ,GaoYT,LouC,ZhangSG,BaiT,WangYJ,SongWQ.
MethylationofDickkopf-3asaprognosticfactorincirrhosis-relatedhepatocellularcarcinoma.
WorldJGastro-enterol2010;16:755-763144SatoH,SuzukiH,ToyotaM,NojimaM,MaruyamaR,SasakiS,TakagiH,SogabeY,SasakiY,IdogawaM,SonodaT,MoriM,ImaiK,TokinoT,ShinomuraY.
Frequentepigeneticinac-tivationofDICKKOPFfamilygenesinhumangastrointesti-naltumors.
Carcinogenesis2007;28:2459-2466145MaehataT,TaniguchiH,YamamotoH,NoshoK,AdachiY,MiyamotoN,MiyamotoC,AkutsuN,YamaokaS,ItohF.
TranscriptionalsilencingofDickkopfgenefamilybyCpGislandhypermethylationinhumangastrointestinalcancer.
WorldJGastroenterol2008;14:2702-2714146HirataH,HinodaY,NakajimaK,KawamotoK,KikunoN,KawakamiK,YamamuraS,UenoK,MajidS,SainiS,IshiiN,DahiyaR.
WntantagonistgeneDKK2isepigeneti-callysilencedandinhibitsrenalcancerprogressionthroughapoptoticandcellcyclepathways.
ClinCancerRes2009;15:5678-5687147DingZ,QianYB,ZhuLX,XiongQR.
PromotermethylationandmRNAexpressionofDKK-3andWIF-1inhepatocellularcarcinoma.
WorldJGastroenterol2009;15:2595-2601148JiangT,HuangL,WangS,ZhangS.
ClinicalsignificanceofserumDkk-3inpatientswithgynecologicalcancer.
JObstetGynaecolRes2010;36:769-773149UrakamiS,ShiinaH,EnokidaH,HirataH,KawamotoK,KawakamiT,KikunoN,TanakaY,MajidS,NakagawaM,IgawaM,DahiyaR.
Wntantagonistfamilygenesasbiomark-ersfordiagnosis,staging,andprognosisofrenalcellcarci-nomausingtumorandserumDNA.
ClinCancerRes2006;12:FatimaSetal.
DKKsandWnt/β-cateninsignalling325August10,2011|Volume2|Issue8|WJCO|www.
wjgnet.
com6989-6997150PineauP,NagaiH,PrigentS,WeiY,GyapayG,WeissenbachJ,TiollaisP,BuendiaMA,DejeanA.
Identificationofthreedistinctregionsofallelicdeletionsontheshortarmofchro-mosome8inhepatocellularcarcinoma.
Oncogene1999;18:3127-3134151HirataH,HinodaY,MajidS,ChenY,ZamanMS,UenoK,NakajimaK,TabatabaiZL,IshiiN,DahiyaR.
DICKKOPF-4activatesthenoncanonicalc-Jun-NH2kinasesignalingpath-waywhileinhibitingtheWnt-canonicalpathwayinhumanrenalcellcarcinoma.
Cancer2011;117:1649-1660152AustinatM,DunschR,WittekindC,TannapfelA,GebhardtR,GaunitzF.
Correlationbetweenbeta-cateninmutationsandexpressionofWnt-signalingtargetgenesinhepatocellu-larcarcinoma.
MolCancer2008;7:21153LepourceletM,ChenYN,FranceDS,WangH,CrewsP,PetersenF,BruseoC,WoodAW,ShivdasaniRA.
Small-mol-eculeantagonistsoftheoncogenicTcf/beta-cateninproteincomplex.
CancerCell2004;5:91-102154EmamiKH,NguyenC,MaH,KimDH,JeongKW,EguchiM,MoonRT,TeoJL,KimHY,MoonSH,HaJR,KahnM.
Asmallmoleculeinhibitorofbeta-catenin/CREB-bindingproteintranscription[corrected].
ProcNatlAcadSciUSA2004;101:12682-12687155ThorneCA,HansonAJ,SchneiderJ,TahinciE,OrtonD,CselenyiCS,JerniganKK,MeyersKC,HangBI,WatersonAG,KimK,MelanconB,GhiduVP,SulikowskiGA,LaFleurB,SalicA,LeeLA,MillerDM,LeeE.
Small-moleculeinhibi-tionofWntsignalingthroughactivationofcaseinkinase1α.
NatChemBiol2010;6:829-836156HeB,YouL,UematsuK,XuZ,LeeAY,MatsangouM,Mc-CormickF,JablonsDM.
AmonoclonalantibodyagainstWnt-1inducesapoptosisinhumancancercells.
Neoplasia2004;6:7-14157YouL,HeB,XuZ,UematsuK,MazieresJ,FujiiN,MikamiI,ReguartN,McIntoshJK,Kashani-SabetM,McCormickF,JablonsDM.
Ananti-Wnt-2monoclonalantibodyinducesapoptosisinmalignantmelanomacellsandinhibitstumorgrowth.
CancerRes2004;64:5385-5389158BehariJ,ZengG,OtrubaW,ThompsonMD,MullerP,Mic-senyiA,SekhonSS,LeoniL,MongaSP.
R-Etodolacdecreasesbeta-cateninlevelsalongwithsurvivalandproliferationofhepatomacells.
JHepatol2007;46:849-857159PrangeW,BreuhahnK,FischerF,ZilkensC,PietschT,Pet-meckyK,EilersR,DienesHP,SchirmacherP.
Beta-cateninaccumulationintheprogressionofhumanhepatocarcino-genesiscorrelateswithlossofE-cadherinandaccumulationofp53,butnotwithexpressionofconventionalWNT-1targetgenes.
JPathol2003;201:250-259160OvejeroC,CavardC,PérianinA,HakvoortT,VermeulenJ,GodardC,FabreM,ChafeyP,SuzukiK,RomagnoloB,YamagoeS,PerretC.
Identificationoftheleukocytecell-derivedchemotaxin2asadirecttargetgeneofbeta-cateninintheliver.
Hepatology2004;40:167-176161CavardC,TerrisB,GrimberG,ChristaL,AudardV,Rad-enen-BussiereB,SimonMT,RenardCA,BuendiaMA,PerretC.
Overexpressionofregeneratingislet-derived1alphaand3alphagenesinhumanprimarylivertumorswithbeta-cateninmutations.
Oncogene2006;25:599-608162MukhopadhyayM,GorivodskyM,ShtromS,GrinbergA,NiehrsC,MorassoMI,WestphalH.
Dkk2playsanessentialroleinthecornealfateoftheocularsurfaceepithelium.
Devel-opment2006;133:2149-2154163LiX,LiuP,LiuW,MayeP,ZhangJ,ZhangY,HurleyM,GuoC,BoskeyA,SunL,HarrisSE,RoweDW,KeHZ,WuD.
Dkk2hasaroleinterminalosteoblastdifferentiationandmineralizedmatrixformation.
NatGenet2005;37:945-952164BarrantesIdelB,Montero-PedrazuelaA,Guadao-FerrazA,ObregonMJ,MartinezdeMenaR,Gailus-DurnerV,FuchsH,FranzTJ,KalaydjievS,KlemptM,HlterS,RathkolbB,Re-inhardC,MorrealedeEscobarG,BernalJ,BuschDH,WurstW,WolfE,SchulzH,ShtromS,GreinerE,HrabédeAngelisM,WestphalH,NiehrsC.
Generationandcharacterizationofdickkopf3mutantmice.
MolCellBiol2006;26:2317-2326S-EditorTianLL-EditorWebsterJRE-EditorZhengXMFatimaSetal.
DKKsandWnt/β-cateninsignalling
- portaljjcao.com相关文档
- Parityjjcao.com
- Praseodymiumjjcao.com
- Kojjcao.com
- 外延jjcao.com
- purposejjcao.com
- importantjjcao.com
瓜云互联-美国洛杉矶高防CN2高防云服务器,新老用户均可9折促销!低至32.4元/月!
瓜云互联一直主打超高性价比的海外vps产品,主要以美国cn2、香港cn2线路为主,100M以内高宽带,非常适合个人使用、企业等等!安全防护体系 弹性灵活,能为提供简单、 高效、智能、快速、低成本的云防护,帮助个人、企业从实现网络攻击防御,同时也承诺产品24H支持退换,不喜欢可以找客服退现,诚信自由交易!官方网站:点击访问瓜云互联官网活动方案:打折优惠策略:新老用户购买服务器统统9折优惠预存返款活动...

美国cera机房 2核4G 19.9元/月 宿主机 E5 2696v2x2 512G
美国特价云服务器 2核4G 19.9元杭州王小玉网络科技有限公司成立于2020是拥有IDC ISP资质的正规公司,这次推荐的美国云服务器也是商家主打产品,有点在于稳定 速度 数据安全。企业级数据安全保障,支持异地灾备,数据安全系数达到了100%安全级别,是国内唯一一家美国云服务器拥有这个安全级别的商家。E5 2696v2x2 2核 4G内存 20G系统盘 10G数据盘 20M带宽 100G流量 1...
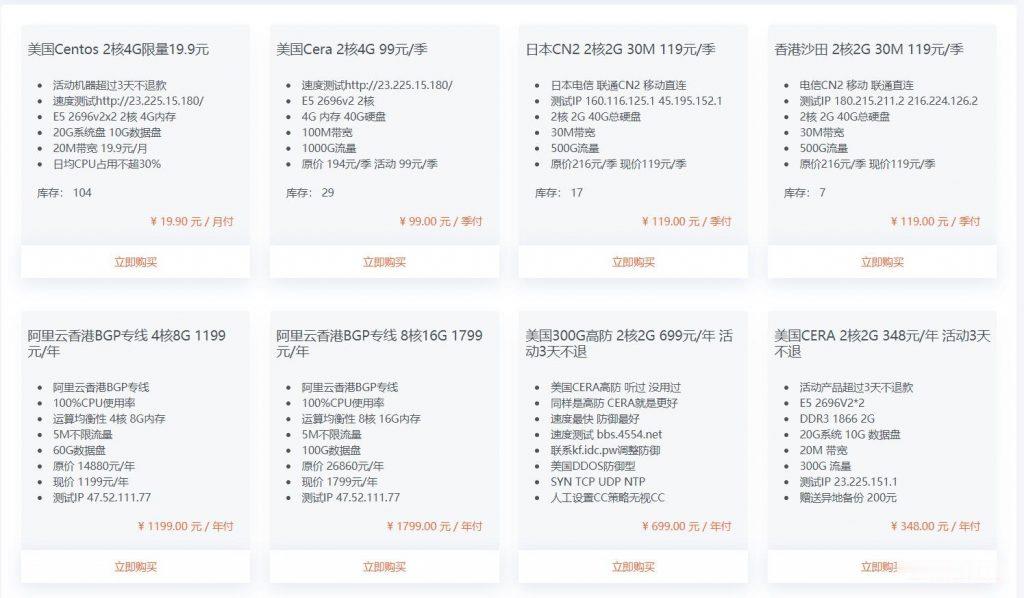
桔子数据58元/月 ,Cera美西云服务器 2核4G 50G数据盘 500M带宽 1000G流量
桔子数据(徐州铭联信息科技有限公司)成立于2020年,是国内领先的互联网业务平台服务提供商。公司专注为用户提供低价高性能云计算产品,致力于云计算应用的易用性开发,并引导云计算在国内普及。目前公司研发以及运营云服务基础设施服务平台(IaaS),面向全球客户提供基于云计算的IT解决方案与客户服务,拥有丰富的国内BGP、双线高防、香港等优质的IDC资源。 公司一直秉承”以人为本、客户为尊、永...

jjcao.com为你推荐
-
巨星prince去世有几位好莱坞巨星死在2016年7788k.comwww.k6320.com 大家给我看看这网站是真是假...rawtoolsRAW是什么衣服牌子巫正刚想在淘宝开一个类似于耐克、阿迪之类的店、需要多少钱、如何能够代理seo优化工具SEO优化要用到什么软件?www.zhiboba.com看NBA直播的网站哪个知道www.diediao.com跪求鸭王2本冈一郎本冈一郎的官网说是日本相扑用的,我们平常的人增肥可以吗?汴京清谈汴京还被称为什么?29ff.comhttp://fcm.com在哪里输入这个网址啊