Priorqqq258.com
qqq258.com 时间:2021-04-09 阅读:()
ORIGINALPAPERAnalysisofintactladderanephospholipids,originatingfromviableanammoxbacteria,usingRP-LC-ESI-MSIngelaLanekoff&RogerKarlssonReceived:4April2010/Revised:3June2010/Accepted:8June2010/Publishedonline:17June2010#TheAuthor(s)2010.
ThisarticleispublishedwithopenaccessatSpringerlink.
comAbstractSincethediscoveryoftheanaerobicammoniumoxidizing(anammox)bacteria,manyattemptshavebeenmadeinordertoidentifytheseenvironmentallyimportantbacteriainnaturalenvironments.
Anammoxbacteriacontainauniqueclassoflipids,calledladderanelipidsandherewepresentanovelmethodtodetectviableanammoxbacteriainsedimentsandwastewatertreatmentplantsbasedontheuseofaladderanelipidbiomarker.
Intactladderanephosphatidylcholine(PC)lipidsareanalyzedusingreversed-phaseliquidchromatography–electrosprayionization–massspectrometry.
Followingextractionfromthecomplexsedimentmatrix,reversed-phaseLCisusedtoseparateladderanePClipidsbasedontheirtailgrouphydrophobicityaswellastheiretheroresterlinktotheglycerolbackboneinthesn-2position.
WeinvestigatethepresenceofintactladderanelipidsinnaturalsedimentsdisplayinganammoxactivityandillustratetheuseofaspecificintactmembraneformingPClipidasabiomarkerforviableanammoxbacterialcells.
Thepresentedmethodcanbeusedtoelucidatethewhereaboutsofviableanammoxbacteria,subsequentlyenablinganestimationofanammoxactivity.
Thiswillgreatlyincreasetheknowledgeofanammoxbacteriaandtheirimportanceintheglobalnitrogencycle.
KeywordsLadderanelipids.
LCMS.
Biomarker.
Anammoxbacteria.
NitrogencycleIntroductionItisestimatedthat25%to50%ofthetotalglobalmarinenitrogengas(N2)productionproceedsviatheanaerobicammoniumoxidation(anammox)reaction[1,2].
Locally,theanammoxcontributionmightbeevenhigher[2].
Bacteriaperforminganammoxwerediscoveredinthe1990sinawastewaterreactorandhavesincebeenfoundinamultitudeofenvironments[3–7].
Anammoxbacteriaconvertbiologi-callyavailablenitrogenspecieslikeammonia,nitrateandnitriteintoN2,whichisreturnedtotheatmosphere.
Inthissensetheyarecrucialforsustainingthenitrogenbalanceinourenvironment[8]andgreateffortshavebeenmadeinthesearchforevidenceoftheirpresenceandactivityinnaturalenvironments.
15N-tracerstudieshavebeenusedtodetermineratesofanammoxbymeasuringtheformationof29N2followingincubationof15N-labelednitrate[9,10].
Togetherwithpredictionsfromnutrientandoxygenprofiles,thismethodhasbeenveryusefulinelucidatingwheretheanammoxreactiontakesplace[11–16].
However,studiesusing15N-labelscannotalonebeusedtostatewhethertheanammoxbacteriaareinvolvedornotsincetheanammoxreactionalsocantakeplacethroughalternativeroutes[17].
Using16SrRNAgeneanalysisandmoleculartechniquessuchasfluorescenceinsituhybridization(FISH)andquantitativepolymerasechainreaction(qPCR),anammoxbacteriahavebeenfoundbothinsedimentsandwaterpillars[7,11,13,18,19].
Furthermore,FISHhasbeenasuccessfulmethodologynotonlyfordeterminingthepresencebutalsothediversityofanammoxbacteriaspecies.
However,FISHisamethodthatreliesonfluorescencemeasurementsthusmakingquantitativemeas-urementsproblematicsincemineralandorganicparticlestendtoquenchthefluorescencesignalwhensedimentI.
Lanekoff:R.
Karlsson(*)DepartmentofChemistry,UniversityofGothenburg,Kemivagen10,41296Gothenburg,Swedene-mail:rogerk@chem.
gu.
seAnalBioanalChem(2010)397:3543–3551DOI10.
1007/s00216-010-3913-3samplesareanalyzed.
ThequantitativedeterminationofanammoxbacterialabundancethroughqPCRisalsoassociatedwithissues.
ObservationshaveshownthatbothRNAandDNAcanberelativelystableaftercelllysis[20,21]resultinginenhancedsignalsduetofossilRNAandDNA[22,23].
Hence,thistechniquemightnotreflecttheactualamountofviableanammoxbacteria.
Ourapproachistodeterminethepresenceofanammoxbacteriabyusingaspecificintactmembraneformingphosphatidylcholinelipidfromthefamilyofso-calledladderanelipidsasabiomarkerforthesebacteriainsediment[2,3,6,7,13,24].
Ofallbacterialcellsexaminedtodate,ladderanelipidshaveonlybeenfoundinanammoxbacteria[25].
Ladderanelipidscontainlinearlyconcatenatedcyclo-butanestructuresattheendofthehydrocarbontails,formingaladder-likestructure(Fig.
1).
Thesehighlystrainedstructuresarefoundasphosphatidylcholine(PC),phospha-tidylethanolamine(PE),andphosphatidylglycerol(PG)lipidsaswellasfattyacids(FA)andlysophospholipids[26–28].
Theintactladderanelipidsexistpredominantlywithanetherlinkagetothesn-2positionofaglycerolbackbone,viaanalkylchain,whilethesn-1positionislinkedviaeitheranetheroresterbond[26,29].
Capillarygaschromatography(GC)massspectrometry(MS)hasbeenusedtoanalyzeladderanephospholipidsasladderaneFAMEs(fattyacidmethylesters)afterderivatization[30].
However,thechromatogramsobtainedsufferedfromverybroadpeaks,containingseveralsmallpeakswithalmostidenticalmassspectra.
Thissuggestedthatthestrainedcyclobutanestruc-tureoftheladderaneFAwasthermallyunstableanddegradedduringGCanalysis[30,31].
AnimprovedmethodfortheanalysisofladderaneFAsbasedonnormal-phasehigh-performanceliquidchromatography(NP-HPLC)–atmosphericpressurechemicalionizationtandemmassspectrometry(APCI-MS/MS)waslaterdeveloped[32],aswellasamethodfortheanalysisofaladderanemonoetherPClysolipid[33].
However,asthesetechniquesrelyontheanalysisofFAsandlysophospholipidstheyhavenotprovidedadirectlinktotheamountoflivinganammoxbacteria.
In2009,Jaeschkeetal.
[33]showedthattheC20-[3]-ladderanemonoetherPC,alysophospholipidretain-inganintactPCheadgroupbuthavingonlyonealkylchain,wasabetterbiomarkerforviableanammoxbacteriathantheladderaneFAs.
However,lysophospholipidsaswellasFAsareonlypresentinverylowamountsinlivingcellssincethemajorityofthemembraneisconstitutedofintactlipidscontainingtwonon-polarhydrocarbontailgroupslinkedtoapolarheadgroup.
Duringdegradation,theintactmembraneformingphospholipidsreadilylosetheirheadgroupbutalsooneofthealkylchains,especiallyifitislinkedtotheglycerolbackboneviaanesterbond,resultinginfattyacidsandlysophospholipidsasdegradationproducts.
Inseveralarticles,ithasbeenshownthatintactmembraneformingphospholipidsisareliabletoolfordetectingviablebacteriainsediments[34–40],duetotherapiddecompositionofphospholipidsaftercelldeath.
EarlierworkperformedbyBoumannetal.
[26]hasdemonstratedtheseparationanddetectionofintactladderanelipidsextractedfrombacterialcellculturesandwastewatertreatmentplants,usingnormal-phaseLCandMSdetection.
ThemethodseparatesintactphospholipidsintoPC,PEandPGspecies,butitdoesnotallowforseparationofthelipidsbasedontheirtailgroupsorbondtotheglycerolbackbone.
Earlierstudiesindicatethattherearemanydifferenttypesofladderanelipidspecies,duetoawiderangeofhydrocarbonchainsbondedtotheglycerolbackboneeitherviaanetheroresterbondinthesn-1position.
Whenaspecificintactlipidistobeusedasabiomarkerforviableanammoxbacteria,itisofutmostimportancethatthemethodofuseseparatesthelipidsintoindividualladderanelipidspecies.
Usingnormal-phaseLCseparation,however,itisimpossibletodistinguishbetweenindividualspecieshavingdifferenttailgroupsbutsharingthesameheadgroup.
ThemethodbyBoumannetal.
[26]hasnotbeenusedforanalyzingintactmembraneformingladderanelipidsfromanammoxbacteriainsedimentsamples.
Theanalysisoflipidsoriginatingfromsedimentsiscomplicatedbythepresenceofmanydifferentphospholipidspeciesaswellasamuchhigheramountofcontaminatingorganicmaterialascomparedtowastewatertreatmentsamples.
ThisputshighdemandsontheLCmethodtoseparateoutspecificindividualphospholipidstobeusedasbiomarkersinsedimentsamples.
Here,wehavedevelopedamethodusingaspecificintactmembraneformingladderanePClipid(m/z816,Fig.
1IV)asabiomarkerforviableanammoxbacterialcellsinnaturalsedimentsdisplayinganammoxactivity.
ThemethodenablesPClipidstobeseparatedbasedontheirtailgrouphydrophobicityaswellastheirattachmenttotheglycerolbackbone,whichcanbeeitherthroughanether–etherorester–etherbond.
ThisenablesanalysisofspecificladderanePClipidsinanammoxbacteriafromnaturaloceansediments.
HereweshowthatthepresenceofthespecificladderanePClipidatdifferentdepthsinasedimentcorecorrelateswellwithestablishednutrientandoxygenprofilesinsedimentsaswellasprofilesofanammoxbacterialabundanceestimatedusingmoleculartechniquesincludingFISHandqPCR[33].
ExperimentalsectionChemicalsPhospholipidreferencesamples1,2-didecanoyl-sn-glycero-3-phosphocholine(10PC),1,2-dipalmitoyl-sn-glycero-3-phospho-choline(16PC),1,2-diphytanoyl-sn-glycero-3-phosphocholine(4Me16PC),1,2-di-O-hexadecyl-sn-glycero-3-phosphocholine3544I.
Lanekoff,R.
Karlsson(DE16PC),and1,2-distearoyl-D70-sn-glycero-3-phosphocho-line(D7018PC)werefromAvantiPolarLipids.
ForLCMSmeasurementsLCMSgrademethanol(MeOH)and2-propanol(isopropylalcohol,IPA)werefromRiedel-deHan,formicacidfromFlukaandammoniafromMerck.
Sodium(meta)periodate,chloroform,n-hexane,ammoniumbicarbonate,anddiethyletherwerefromSigma-Aldrich.
Dichloromethane(DCM)anddichlorodimethylsilanewerefromFlukaandNH2solid-phaseextraction(SPE)columns(500mg,3mL)fromStrata.
MaterialsFormethoddevelopment,anammoxbacteriagrowinginbiofilmsonplasticbiofilmcarriers(AnoxkaldnesAB,Lund,Sweden)wereobtainedfromHimmerfjrdsverketswastewatertreatmentplantinStockholm,Sweden.
LadderanePClipidsextractedfromanammoxbacteriabiofilmswereusedasreferencestodeterminetheLCretentiontimeduringsedimentanalysis.
Sedimentcoresamples(dividedintodepths0–3,3–6,6–9,and9–12cm)wereobtainedfromGullmarsfjordenoutsideLysekilonthewestcoastofSwedenandstoredat4°Cforafewdaysuntilanalysiswasperformed.
BiofilmextractionToextractladderanelipids,threedifferentprotocolswereinvestigated.
ThefirstprotocolisdescribedbyZinkandMangelsdorf[41]whereMeOH:DCM:10mMammoniumacetate(AmAc)isusedintheratioof2:1:0.
8.
ThesecondfollowstheFolchprocedureusingamixtureofchloroform:MeOH(2:1)[42]andthethird,fromHaraetal.
[43],usesamixturebasedonn-hexane:isopropanol(3:2).
Priortoextractionthebiofilmsampleswerecentrifugedandcarefullyweighedtoensurethatacomparisonbetweenthedifferentlytreatedbiofilmsamplescouldbemade.
Inallmethodsweusedultrasonication,asdescribedbelow,andextractedthelipidsthreetimeswithacentrifugationstepinbetween.
Supernatantswerecombinedandcollectedinseparateseparationfunnels.
Allbiofilmsamplesweresubjectedtoliquid–liquidextraction(LLE),asdescribedintheoriginalpapers[41–43],andSPE[44].
Thephospholipidfractionswerefullyevaporatedinasilanizedtesttubeunderastreamofnitrogengas,re-dissolvedin200μLmobilephase,andstoredforashortperiodoftimeundernitrogenatmosphereat80°CpriortoLCMSanalysis.
TheFolch[42]andHara[43]procedureswerelessefficientthantheZinkprocedure[41]andwerethusnotusedforthesedimentextractions.
SedimentincubationandextractionAsedimentcorewasobtainedfromGullmarsfjorden(110mbelowseasurface)anddividedintofour3-cm-thicksamplesbetween0and12cmofdepth.
Anaqueoussolutioncontaining5%sodium(meta)periodate(150ml)wasaddedto150gofwetsediment.
Thesedimentslurrywassonicatedfor2min,usinganultrasonicprocessor(SonicsVibracell,Model501)at70%,witha7-spulseanda5-srest.
Theslurrywasincubatedat4°Cforaperiodof1-weektobreakclustersofbacteriaandtolysecells[45,46].
Afterincubation,thesamplewastransferredtotwo250-mLcentrifugeflasks(TeflonFEP,Nalgene)andcentrifugedat3,220gfor15minusinganEppendorfCentrifuge5810R.
Thesupernatantswerecombinedandcollectedinaseparationfunnel.
MeOH(75ml)wasaddedtoeachcentrifugeflaskandthesamplewassonicatedandcentrifugedusingthesameconditionsasabove.
TheMeOHsupernatantswerealsocombinedandcollectedinaseparateseparationfunnel.
Tofurtherextractlipids,theZinkprocedure[41]wasused.
Thesamplewasagaintreatedwithultrasonicationasdescribedabovebutwith75mLofMeOH:DCM:AmAc(2:1:0.
8)threetimesperflaskwithcentrifugationstepsinbetween[41].
Thesupernatantswerecombinedandcollectedinseparateseparationfunnels.
Fig.
1SchematicillustrationsofladderanePClipidsinvestigatedinthisstudy.
(I)Dietherlipidatm/z800,(II)dietherlipidatm/z802,(III)ether/esterlipidatm/z814,(IV)ether/esterlipidatm/z816.
Notethesequentiallylinkedcyclobutane/cyclohexanestructuresattheendofthehydrocarbontailsIntactladderanephospholipidsfromviableanammoxbacteria3545SedimentsamplecleanupprocedurePriortoLLE,DCM,andAmAcwereaddedtoalltheseparationfunnelsuntiltheratiosofthesolventswereMeOH:DCM:AmAc1:1:0.
9.
Afterphaseseparation,theDCMphaseswerecollectedinaroundbottomflask.
ThewaterphasewassubjectedtotwomoreextractionswhereDCM:MeOHwasaddedinaratioof2:1[41].
Again,theDCMphaseswerecollectedandcombinedwiththepreviousextracts.
TheDCMintheroundbottomflaskwasreducedusingarotavaporatorfromBüchi,andfullyevaporatedinasilanizedtesttubeunderaflowofnitrogengas.
Thesamplewasre-dissolvedin12mLof2:1n-hexane:chloroformandstoredundernitrogenatmo-sphereat80°CuntilsubjectedtofurthercleanupbySPE.
FourSPEcolumns(StrataNH2(55μm,70A)500mg,3mL)persamplewereconnectedtoavacuumbox(Resprepvacuummanifoldwith12ports)andactivatedwith12mLMeOH.
Thecolumnswerecondi-tionedwith12mLofn-hexanepriortoloadingthesample.
Awashwith12mLchloroform:IPA(2:1)elutedcholesterylesters,triglycerides,cholesterols,diglycerides,andmonoglycerides.
FAswereelutedwith2%aceticacidindiethylether.
Finally,thephospholipidswereelutedwith12mLofMeOH,combinedandfullyevaporatedinasilanizedtesttubeunderastreamofnitrogengas[44].
Thecleansamplewasre-dissolvedin200μLmobilephase,andstoredundernitrogenatmosphereforashortperiodoftimeat80°CpriortoLCMSanalysis.
LiquidchromatographyandmassspectrometrySeparationwasachievedusingaGeminiC18column(50*2mm,3μmparticlediameter,hybridsilicagelcolumnforreversed-phaseHPLC)withaGeminiC18guardcolumn(4*2mm),fromPhenomenex.
Thetemper-atureofthecolumnwaskeptconstantat30°CusingacolumnovenLCO102fromEcom.
ThemobilephasesinthegradientconsistedofA;4%water,0.
1%formicacid,and0.
1%ammonia(25%)inMeOHandB;isopropanol(IPA)containing0.
1%formicacidand0.
1%ammonia(25%).
Theinitialcompositionwithaflowof0.
4mL/minwas100%Awhichwasloweredto70%Abetween10and20minoftheseparation.
At21min,theflowwasloweredto0.
2mL/minand100%Bwasusedtowashthecolumnfor10minbeforereturningtotheinitialconditions.
Totalanalysistime,includingwashingstep,foreachsamplewas40min.
Theanalyteswereelutedwithin20min.
Analysisofbiofilmsamplesduringmethoddevelopmentwasperformedusinga600SControllerand626PumpfromWatersanda234autosamplerfromGilsoncoupledtoaMicromassQuattroLC,triplequadrupole,withelectrosprayionization(ESI).
MS/MSanalysisenabledparentalionscanningofthePCheadgroup,atm/z184,todetectPClipids.
Argonwasusedascollisiongas,thepotentialoverthecapillarywas4.
0kVandtheconevoltagewas90V.
Sourceblocktemperaturewasheldat150°Canddesolvationtemperatureat300°C.
SedimentsampleanalysiswasperformedusingaPerkin-ElmerSeries200autosamplerandLCpumpcoupledtoaMicromassQToF,quadrupoletime-of-flightwithESIscanningbetweenm/z100and900.
AsaretentiontimereferenceforladderanePClipidsabiofilmsampleextractwasconsecutivelyanalyzedusingtheQToFset-up.
ResultsanddiscussionLadderanePClipidextractionandsamplecleanupAnammoxbacteriahavebeenshowntoformbiofilmswhenculturedinwastewaterreactorsandmostlikelygrowinclustersinoceansedimentsaswell.
Biofilmandclustersaremostlyformedviaaprotein-polysaccharidematrix[47,48]andbiofilmformationmakesitmoredifficulttolysecellsinordertoextractlipidsfromthecellmembranes.
Forthisstudy,severalchemicalandphysicaltreatments,includingboiling,hydrogenperoxideorhypochloratetreatments,mechanicalhomogenizationandsonicationwereattemptedinordertoseparateandlysecellsinbiofilms(datanotshown).
Thesuccessofthesetreatmentsweremonitoredusingfluorescencemicroscopybystainingthecellswith4′,6-diamidino-2-phenylindole.
Furthermore,theextractedladderanePClipidswereanalyzedusingLCMSpostextraction.
Thebestresults,showedbylackofladderanelipidsinsamplesextractedasecondtime,wereachievedwhenanammoxbacteriainbiofilmswereincubatedwitha5%solutionofperiodatepriortoprobesonication,asdescribedintheexperimentalsection[45,46].
NosignificantdifferenceintheladderanePClipidcompositionorquantitydependingontheperiodateincubationtime(2daysversus7days)wereobserved,showingthatnodecompositionoftheladderanePClipidoccurredduringincubation(datanotshown).
InoceansedimentsfromGullmarsfjorden,approximately3%ofallbacteriahavebeenfoundtobeanammoxbacteria[4].
Thelowcellabundanceandthecomplexityofthesedimentsamplematrixputshighdemandsontheextractionmethod.
TooptimizetheyieldofladderanePClipidsintheextractionstep,wecomparedthreeextractionprotocols,usingcarefullyweighedtriplesamplesofanammoxbacteriainbiofilms.
Thesolventmixturesinthecomparedmethodswere;chloroform:MeOH[42],n-hexane:isopropanol[43]andMeOH:DCM:AmAc[41].
Afterextractionandsamplecleanup,allsampleswereanalyzedusingthedescribedLCMSmethod.
InFig.
2,theareaoftheintegratedpeaksof3546I.
Lanekoff,R.
KarlssonthreechosenladderanePClipids,foundatm/z802,814,and816,arecompared.
TheresultshowsthatthehighestyieldofladderanePClipidsisobtainedwhenthemixtureofMeOH:DCM:AmAc[41]isused.
LCmethodforladderanelipidsThedevelopedreversed-phaseLCmethodseparatesPClipidsbasedonthetailgrouphydrophobicity,makingtheladderanelipidselutelast.
Figure3bshowsthechromato-gramgeneratedduringseparationofthefivenon-ladderanePClipidstandardsshowninFig.
3a.
ThisconfirmsthattheLCmethodseparatesthePClipidsbasedonthelengthandhydrophobicityofthehydrocarbontailgroup.
Furthermore,themethodseparatesthePClipidsbasedonthelinktotheglycerolbond,wherethemorepolarether–ester-linkedlipidselutebeforetheether–etherlinkedlipidswiththesametailgroups.
Thisfeatureoftheseparationisespeciallyimportantwhenaspecificladderanelipidisbeingusedasabiomarker.
TheconcentrationsofalllipidsinFig.
3barethesame,buttheresponsevariesconsiderably.
Themostobviousdifferenceisthattheresponseoftheether–etherlipidisseveralmagnitudeslower.
ThisdecreasedsignalintensitycouldbeduetothelowerionizationorfragmentationefficiencyofthedietherPCduringmassspectrometricanalysis[49].
Thehigherdetectionlimitofdietherspeciesencouragedtheuseofanondietherspecieasbiomarker.
InFig.
3c,abiofilmsamplecontaininganammoxbacteriaisanalyzedusingparentalionscanningofthePCheadgroup.
Fig.
3ChromatogramsshowingtheLCseparationofladderanePClipids.
aStructuresoffivenon-ladderanePClipidsusedduringmethoddevelopment.
(1)10PC,(2)16PC,(3)DE16PC,(4)D7018PC,(5)4Me16PC.
bChromatogramofthenon-ladderanePCstandardsatthesameconcentrationanalyzedbyRP-LCQqQusingparentalionscanningofm/z184.
cChromatogramofabiofilmsamplecontaininganammoxbacteriaanalyzedbyRP-LCQqQusingparentalionscanningofm/z184.
Ladderanelipidselutelastinthechromatogram.
Themorepolarether–ester-linkedladderanePClipidsIIIandIVelutebeforetheether–etherlinkedIandIIFig.
2ComparisonofthreeextractionmethodsforladderanePClipidsfoundatm/z802(II),814(III),and816(IV),n=3.
TheRomannumeralsII,III,andIVdenotethestructuresillustratedinFig.
1.
*Denotesasignificantdifference(studentsttest<0.
05)bIntactladderanephospholipidsfromviableanammoxbacteria3547ThealmostidenticalladderanePClipidsatm/z814(III)andm/z816(IV),bothbeingester–etherladderanePClipids,coeluteinthepeakdenotedbyIII,IVinFig.
3c.
Asexpected,theladderanePClipidsatm/z800(I)andm/z802(II),alsoalmostidenticalbutbothdietherladderanePClipids,coelutelaterinthepeakdenotedbyI,II.
ThecoelutionofspeciesIIIandIVaswellasIandIIisacceptablesincetheseanalytesoriginatefromtheanammoxbacteriainconstantrelativeconcentrations,thuskeepinganeventualdiscriminationeffectduringanalysisconstant.
MassspectrometryofladderanelipidsForelectrospraymassspectrometry,PClipidsionizebestinthepositiveionmodeandreadilylosetheirheadgroupatm/z184whensubjectedtocollisioninduceddissociation.
AQqQ,usingparentalionscanningofthem/z184fragmention,wasusedforanalyteconfirmationduringmethoddevelopment.
Figure4ashowsthechromatogramofananammoxbiofilmsample.
ThetwopeakselutinglastcontaintheintactladderanePClipidsI–IVasshowninthecorrespondingmassspectra(Fig.
4b–c).
ThedetectionlimitsoftheladderanePClipidswasfoundtobehigherwhenusingparentalionscanningontheQqQcomparedtoscanningusingtheQToF.
ThiscanpartlybeexplainedbythefactthattheladderanePClipidshavecomparablylonghydrocarbontailgroupswhichmakethemhardtofragment[46].
Therefore,weusedtheQToF,scanningbetweenm/z100and900,todetectladderanePClipidsinsedimentsampleswhichcontainsubstantiallyloweramountsthanbiofilmsamples.
InFig.
4d,asedimentsampleisanalyzedbyRP-LCQToFandtheselectedmasschromatogram,i.
e.
achromatogramwherethem/z816,correspondingtotheladderanePClipidIV,isextractedoutofthetotalionchromatogram,isshown.
ThepeaklabeledasIVistheladderanePClipidatm/z816andthecorrespondingmassspectrumofthatpeakisfoundinFig.
4e.
ThisclearlyshowsthepresenceofanammoxbacteriaintheoceansedimentsamplethatwastakenfromGullmarsfjordenonthewestcoastofSweden.
AscanbeseeninthemassspectrafrombothbiofilmandsedimentsamplestheRP-LCmethodclearlyseparatetheladderanePClipidsfromotherlipidspeciesinthecomplexsedimentsample.
Fig.
4ChromatogramsandmassspectraofintactladderanePClipids.
aChromatogramofabiofilmsamplecontaininganammoxbacteriaanalyzedbyRP-LCQqQusingparentalionscanningofm/z184.
ThetwopeakselutinglateinthechromatogramcontainstheladderanelipidsI–IVasshowninbandc.
bMassspectrumextractedfromthepeakat12.
5mindenotedIII,IVina.
TheintactladderanePClipidsIIIandIVatm/z814and816areidentified.
cMassspectrumextractedfromthepeakat16mindenotedI,IIinA.
TheintactladderanePClipidsIandIIatm/z800and802areidentified.
dSelectedmasschromatogramfortheintactladderanePClipidIVatm/z816inasedimentsampleanalyzedbyRP-LCQToFscanningbetweenm/z100and900.
eMassspectrumextractedfromthepeakat16mindenotedIVind.
TheintactladderanePClipidsIIIandIVatm/z814and816areidentifiedb3548I.
Lanekoff,R.
KarlssonDeterminationofaspecificladderanebiomarkerTheladderanePClipidatm/z816waschosenasabiomarkerforanammoxbacteriabasedontwoimportantcharacteristics.
First;theladderanePClipidatm/z816displaysthehighestabundanceinthemassspectrumwhenusingthedescribedmethod.
Second,theladderanePClipidatm/z816isanether–ester-linkedladderanelipid,whichwehaveobservedtobelessstableinsedimentwhencomparedtotheether–etherlinkedPClipids.
Thelowstabilityinsedimentenvironmentalsosuggeststhatthislipidwouldbettertracklivinganammoxbacteria.
Thiswasinvestigatedbyanalyzingtheamountofester–ether(e.
g.
,m/z816)andether–ether(e.
g.
,m/z800)linkedladderanePClipidsinfreshsedimentandagainafterstoringthatsamesedimentformorethan1monthinarefrigeratorandat20°C.
Theether–esterladderanePClipidswerefoundinreducedlevelsintheoldersedimentscomparedtothefresh,whereastheether–etherladderanePClipidswerefoundinthesameamountsregardlessofsedimenttype.
Thisobservationsuggeststhatthedegradationinsedimentfortheselipidsdoesnotoccuratsimilarratesandthatanester–etherlinkedladderanePClipidisasuitablebiomark-erforlivinganammoxbacteria.
Furthermore,thestabilityoftheether–etherladderanePClipidsoffersanexplanationtowhyotherstudies,analyzingladderanelipiddegradationproducts,havefoundladderaneFAsandladderanelysoli-pidsspreadthroughoutthesedimentdepthprofiles[33].
TodemonstratetheaccuracyoftheladderanePClipidatm/z816asbiomarkerforviableanammoxbacteriawehaveperformedtestsonbothsamplesfromthewastewatertreatmentplantHimmerfjrdsverketinStockholm,SwedenandasedimentcorefromGullmarsfjordenonthewestcoastofSweden.
BiofilmsamplesfromthreeofthereactorsatHimmerfjrdsverketpresumablycontaininganammoxbacteriawereanalyzed.
Thebiomarkerwaspresentinthesamplefromthereactorthatdisplayedanammoxactivity(datanotshown),shownbythegenerationofnitrogengasunderanoxicconditions,indicatingthepresenceofintactlivinganammoxbacteriainthereactor.
DetectingladderanePClipidsinthecomplexmatrixofsedimentisconsiderablymoredifficult.
Thecorrespondingmasschromatogramsofm/z816atthedepths0–3,3–6,6–9,and9–12cmareshowninFig.
5a.
Nutrientandoxygenprofiles,previouslyinvestigatedby,e.
g.
,Jaeschkeetal.
[33],indicatesthatanammoxbacteriaresideintheupperlayerofthesediment,closetotheborderbetweentheanoxicandoxiczone.
Accordingly,theladderanePClipidbiomarkeratm/z816wasfoundatthisdepth,asshowninthedepthprofileinFig.
5b.
Themorestableether–etherladderanePClipidatm/z800was,however,foundatalmosteverydepthinthecore,likelyduetotheslowerdegradationrateforether–etherPClipidsrelativetotheether–esterlipids,asdiscussedabove.
TheamountofladderanePClipidsfoundatthesedepthscanberelatedtoeachothersincetheladderanePClipidshavealinearresponse(integratedpeakareaversusconcentration)intheconcentrationrangeofinterest(datanotshown).
Inthis,examplethedepthsinthecorearerough,andthedepthatwhichtheanammoxbacteriaarefocusedisprobablyalotFig.
5Resultfromthesedimentdepthprofilestudy.
aFouroverlayedmasschromatogramsofm/z816.
Thesedimentsampledepthsareindicatedatthetraces.
ThebottomtraceisareferencesamplefromabiofilmcontainingladderanePClipids.
bDepthprofileofladderanePClipidIVatm/z816(ether–ester),indicatedbythetriangles,andladderanePClipidIatm/z800(ether–ether),indicatedbythecircles.
Depthslicesare3cmthick.
ForladderanePClipidIV,atm/z816,thepeakswerenormalizedtothemaximumareafoundatdepth0–3cmdenoted100%.
ForladderanePClipidI,atm/z800,thepeakswerenormalizedtothemaximumareafoundatdepth6–9cmdenoted100%Intactladderanephospholipidsfromviableanammoxbacteria3549narrower.
ThefindingofanammoxbacteriaatthissiteinGullmarsfjordenisconsistentwithpreviousobservations[4,50].
ConclusionsAmethodhasbeendevelopedtoanalyzeintactladderanePClipidsusingRP-LC-ESI-MS.
Inthisstudy,themethodwasusedtoidentifyaspecificintactladderanePClipidbiomarkerforviableanammoxbacteria.
Thechosenbiomarkerisanether–esterladderanePClipidwhichisfoundatm/z816.
Further,threedifferentmethodsforextractingladderanePClipidshavebeenevaluatedandamultistepsamplecleanupprocedureforladderanelipidsinsedimentsamplesisdescribed.
Whenusingthedescribedmethodonsedimentsamples,wefoundthattheselectedbiomarkerispresentonlyintheupperpartofthesedimentcore.
Thisfindingissupportedbyearlierobservationsthatanammoxbacteriaresidesintheupperpartofthesediment,justbelowtheoxiczone.
Thedescribedmethodprovidesanewtoolfortheanammox-interestedresearchcommunitytostudyviableanammoxbacteriainnaturalenvironments.
Thiswillleadtoanincreasedunderstandingofthepreferredbiogeochemicalenvironmentoftheanammoxbacteriaandhowtheircommunitystructureisaffectedbyongoingenvironmentalchanges.
Furthermore,thelocaliza-tionofviableanammoxbacteriaandaconcomitantobservationofanammoxactivitywillcontributetoagreaterunderstandingoftheglobalnitrogencycle.
AcknowledgmentsTheauthorswouldliketogreatlyacknowledgeProfessorStefanHulthandhisgroupattheDepartmentofChemistry,UniversityofGothenburgforfruitfuldiscussionsaswellashelpwithsamplecollection.
OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttributionNoncommercialLicensewhichper-mitsanynoncommercialuse,distribution,andreproductioninanymedium,providedtheoriginalauthor(s)andsourcearecredited.
References1.
DevolAH(2003)Nitrogencycle—solutiontoamarinemystery.
Nature422:575–5762.
KuypersMMM,LavikG,WoebkenD,SchmidM,FuchsBM,AmannR,JorgensenBB,JettenMSM(2005)MassivenitrogenlossfromtheBenguelaupwellingsystemthroughanaerobicammoniumoxidation.
ProcNatlAcadSciUSA102:6478–64833.
JaeschkeA,denCampH,HarhangiH,KlimiukA,HopmansEC,JettenMSM,SchoutenS,DamsteJSS(2009)16SrRNAgeneandlipidbiomarkerevidenceforanaerobicammonium-oxidizingbacteria(anammox)inCaliforniaandNevadahotsprings.
FEMSMicrobiolEcol67:343–3504.
SchmidMC,Risgaard-PetersenN,vandeVossenbergJ,KuypersMMM,LavikG,PetersenJ,HulthS,ThamdrupB,CanfieldD,DalsgaardT,RysgaardS,SejrMK,StrousM,denCampH,JettenMSM(2007)Anaerobicammonium-oxidizingbacteriainmarineenvironments:widespreadoccurrencebutlowdiversity.
EnvironMicrobiol9:1476–14845.
StrousM,FuerstJA,KramerEHM,LogemannS,MuyzerG,vandePas-SchoonenKT,WebbR,KuenenJG,JettenMSM(1999)Missinglithotrophidentifiedasnewplanctomycete.
Nature400:446–4496.
KuypersMMM,SliekersAO,LavikG,SchmidM,JorgensenBB,KuenenJG,DamsteJSS,StrousM,JettenMSM(2003)AnaerobicammoniumoxidationbyanammoxbacteriaintheBlackSea.
Nature422:608–6117.
SchmidMC,MaasB,DapenaA,dePas-SchoonenKV,deVossenbergJV,KartalB,vanNiftrikL,SchmidtI,CirpusI,KuenenJG,WagnerM,DamsteJSS,KuypersM,RevsbechNP,MendezR,JettenMSM,StrousM(2005)Biomarkersforinsitudetectionofanaerobicammonium-oxidizing(anammox)bacteria.
ApplEnvironMicrobiol71:1677–16848.
KuenenJG(2008)Anammoxbacteria:fromdiscoverytoapplication.
NatRevMicrobiol6:320–3269.
ThamdrupB,DalsgaardT(2002)ProductionofN-2throughanaerobicammoniumoxidationcoupledtonitratereductioninmarinesediments.
ApplEnvironMicrobiol68:1312–131810.
WakiM,YasudaT,SuzukiK,SakaiT,SuzukiN,SuzukiR,MatsubaK,YokoyamaH,OginoA,TanakaY,UedaS,TakeuchiM,YamagishiT,SuwaY(2010)Ratedeterminationanddistributionofanammoxactivityinactivatedsludgetreatingswinewastewater.
BioresourceTechnology101:2685–269011.
DalsgaardT,ThamdrupB,CanfieldDE(2005)Anaerobicammoniumoxidation(anammox)inthemarineenvironment.
ResMicrobiol156:457–46412.
DalsgaardT,CanfieldDE,PetersenJ,ThamdrupB,Acuna-GonzalezJ(2003)N-2productionbytheanammoxreactionintheanoxicwatercolumnofGolfoDulce,CostaRica.
Nature422:606–60813.
SchubertCJ,Durisch-KaiserE,WehrliB,ThamdrupB,LamP,KuypersMMM(2006)Anaerobicammoniumoxidationinatropicalfreshwatersystem(LakeTanganyika).
EnvironMicrobiol8:1857–186314.
TrimmerM,NichollsJC,DeflandreB(2003)Anaerobicammo-niumoxidationmeasuredinsedimentsalongtheThamesestuary,UnitedKingdom.
ApplEnvironMicrobiol69:6447–645415.
RysgaardS,GludRN(2004)AnaerobicN-2productioninArcticseaice.
LimnolOceanogr49:86–9416.
RysgaardS,GludRN,Risgaard-PetersenN,DalsgaardT(2004)DenitrificationandanammoxactivityinArcticmarinesediments.
LimnolOceanogr49:1493–150217.
HulthS,AllerRC,GilbertF(1999)Coupledanoxicnitrificationmanganesereductioninmarinesediments.
GeochimCosmochimActa63:49–6618.
RichJJ,DaleOR,SongB,WardBB(2008)Anaerobicammoniumoxidation(Anammox)inChesapeakeBaysediments.
MicrobEcol55:311–32019.
DaleOR,TobiasCR,SongBK(2009)Biogeographicaldistributionofdiverseanaerobicammoniumoxidizing(anammox)bacteriainCapeFearRiverEstuary.
EnvironMicrobiol11:1194–120720.
NannipieriP,CiardiC,BadaluccoL,CasellaS(1986)AmethodtodeterminesoilDNAandRNA.
SoilBiolBiochem18:275–28121.
SchmidM,TwachtmannU,KleinM,StrousM,JuretschkoS,JettenM,MetzgerJW,SchleiferKH,WagnerM(2000)Molecularevidenceforgenusleveldiversityofbacteriacapableofcatalyzinganaerobicammoniumoxidation.
SystApplMicrobiol23:93–10622.
CoolenMJL,OvermannJ(1998)Analysisofsubfossilmolecularremainsofpurplesulfurbacteriainalakesediment.
ApplEnvironMicrobiol64:4513–45213550I.
Lanekoff,R.
Karlsson23.
CoolenMJL,MuyzerG,RijpstraWIC,SchoutenS,VolkmanJK,DamsteJSS(2004)CombinedDNAandlipidanalysesofsedimentsrevealchangesinHolocenehaptophyteanddiatompopulationsinanAntarcticlake.
EarthPlanetSciLett223:225–23924.
ByrneN,StrousM,CrepeauV,KartalB,BirrienJL,SchmidM,LesongeurF,SchoutenS,JaeschkeA,JettenM,PrieurD,GodfroyA(2009)Presenceandactivityofanaerobicammonium-oxidizingbacteriaatdeep-seahydrothermalvents.
ISMEJ3:117–12325.
JettenMSM,vanNiftrikL,StrousM,KartalB,KeltjensJT,OpdenCampHJM(2009)Biochemistryandmolecularbiologyofanammoxbacteria.
CritRevBiochemMolBiol44:65–8426.
BoumannHA,HopmansEC,vandeLeemputI,OpdenCampHJM,vandeVossenbergJ,StrousM,JettenMSM,DamsteJSS,SchoutenS(2006)Ladderanephospholipidsinanammoxbacteriacomprisephosphocholineandphosphoethanolamineheadgroups.
FEMSMicrobiolLett258:297–30427.
RattrayJE,vandeVossenbergJ,HopmansEC,KartalB,vanNiftrikL,RijpstraWIC,StrousM,JettenMSM,SchoutenS,DamsteJSS(2008)Ladderanelipiddistributioninfourgeneraofanammoxbacteria.
ArchMicrobiol190:51–6628.
BoumannHA,LongoML,StroeveP,PoolmanB,HopmansEC,StuartMCA,DamsteJSS,SchoutenS(2009)Biophysicalpropertiesofmembranelipidsofanammoxbacteria:I.
Ladderanephospholipidsformhighlyorganizedfluidmembranes.
BiochimBiophysActaBiomembr1788:1444–145129.
Sinninghe-DamstéJSS,StrousM,RijpstraWIC,HopmansEC,GeenevasenJAJ,vanDuinACT,vanNiftrikLA,JettenMSM(2002)Linearlyconcatenatedcyclobutanelipidsformadensebacterialmembrane.
Nature419:708–71230.
Sinninghe-DamstéJSS,RijpstraWIC,GeenevasenJAJ,StrousM,JettenMSM(2005)Structuralidentificationofladderaneandothermembranelipidsofplanctomycetescapableofanaerobicammoniumoxidation(anammox).
FEBSJ272:4270–428331.
JaeschkeA,LewanMD,HopmansEC,SchoutenS,DamsteJSS(2008)Thermalstabilityofladderanelipidsasdeterminedbyhydrouspyrolysis.
OrgGeochem39:1735–174132.
HopmansEC,KienhuisMVM,RattrayJE,JaeschkeA,SchoutenS,DamsteJSS(2006)Improvedanalysisofladderanelipidsinbiomassandsedimentsusinghigh-performanceliquidchroma-tography/atmosphericpressurechemicalionizationtandemmassspectrometry.
RapidCommunMassSpectrom20:2099–210333.
JaeschkeA,RooksC,TrimmerM,NichollsJC,HopmansEC,SchoutenS,DamsteJSS(2009)Comparisonofladderanephospholipidandcorelipidsasindicatorsforanaerobicammoni-umoxidation(anammox)inmarinesediments.
GeochimCosmo-chimActa73:2077–208834.
AriesE,DoumenqP,ArtaudJ,MolinetJ,BertrandJC(2001)Occurrenceoffattyacidslinkedtonon-phospholipidcompoundsinthepolarfractionofamarinesedimentaryextractfromCarteaucove,France.
OrgGeochem32:193–19735.
SturtHF,SummonsRE,SmithK,ElvertM,HinrichsKU(2004)Intactpolarmembranelipidsinprokaryotesandsedimentsdecipheredbyhigh-performanceliquidchromatography/electro-sprayionizationmultistagemassspectrometry—newbiomarkersforbiogeochemistryandmicrobialecology.
RapidCommunMassSpectrom18:617–62836.
ZinkKG,WilkesH,DiskoU,ElvertM,HorsfieldB(2003)Intactphospholipids—microbial"lifemarkers"inmarinedeepsubsur-facesediments.
OrgGeochem34:755–76937.
MazzellaN,MolinetJ,SyaktiAD,BertrandJC,DoumenqP(2007)Assessmentoftheeffectsofhydrocarboncontaminationonthesedimentarybacterialcommunitiesanddeterminationofthepolarlipidfractionpurity:relevanceofintactphospholipidanalysis.
MarChem103:304–31738.
H.
Ruetters,H.
Sass,H.
Cypionka,J.
Rullkotter,in4thInternationalSymposiumontheInterfacebetweenAnalyticalChemistryandMicrobiology,Bretagne,France,2000,pp.
149–16039.
ZinkKG,MangelsdorfK,GraninaL,HorsfieldB(2008)Estimationofbacterialbiomassinsubsurfacesedimentsbyquantifyingintactmembranephospholipids.
AnalBioanalChem390:885–89640.
RuettersH,SassH,CypionkaH,RullkotterJ(2002)MicrobialcommunitiesinaWaddenSeasedimentcore—cluesfromanalysesofintactglyceridelipids,andreleasedfattyacids.
OrgGeochem33:803–81641.
ZinkKG,MangelsdorfK(2004)EfficientandrapidmethodforextractionofintactphospholipidsfromsedimentscombinedwithmolecularstructureelucidationusingLC-ESI-MS-MSanalysis.
AnalBioanalChem380:798–81242.
FolchJ,LeesM,StanleyGHS(1957)Asimplemethodfortheisolationandpurificationoftotallipidsfromanimaltissues.
JBiolChem226:497–50943.
HaraA,RadinNS(1978)Lipidextractionoftissueswithalow-toxicitysolvent.
AnalBiochem90:420–42644.
KaluznyMA,DuncanLA,MerrittMV,EppsDE(1985)Rapidseparationoflipidclassesinhigh-yieldandpurityusingbondedphasecolumns.
JLipidRes26:135–14045.
HuebnerJ,WangY,KruegerWA,MadoffLC,MartirosianG,BoisotS,GoldmannDA,KasperDL,TzianabosAO,PierGB(1999)IsolationandchemicalcharacterizationofacapsularpolysaccharideantigensharedbyclinicalisolatesofEnterococcusfaecalisandvancomycin-resistantEnterococcusfaecium.
InfectImmun67:1213–121946.
LiuCH,LeeSM,VanLareJM,KasperDL,MazmanianSK(2008)Regulationofsurfacearchitecturebysymbioticbacteriamediateshostcolonization.
ProcNatlAcadSciUSA105:3951–395647.
ElversKT,LeemingK,MooreCP,Lappin-ScottHM(1998)Bacterial-fungalbiofilmsinflowingwaterphoto-processingtanks.
JApplMicrobiol84:607–61848.
O'TooleG,KaplanHB,KolterR(2000)Biofilmformationasmicrobialdevelopment.
AnnuRevMicrobiol54:49–7949.
Al-SaadKA,SiemsWF,HillHH,ZabrouskovV,KnowlesNR(2003)Structuralanalysisofphosphatidylcholinesbypost-sourcedecaymatrix-assistedlaserdesorption/ionizationtime-of-flightmassspectrometry.
JAmSocMassSpectrom14:373–38250.
vandeVossenbergJ,RattrayJE,GeertsW,KartalB,vanNiftrikL,vanDonselaarEG,DamsteJSS,StrousM,JettenMSM(2008)Enrichmentandcharacterizationofmarineanammoxbacteriaassociatedwithglobalnitrogengasproduction.
EnvironMicrobiol10:3120–3129Intactladderanephospholipidsfromviableanammoxbacteria3551
ThisarticleispublishedwithopenaccessatSpringerlink.
comAbstractSincethediscoveryoftheanaerobicammoniumoxidizing(anammox)bacteria,manyattemptshavebeenmadeinordertoidentifytheseenvironmentallyimportantbacteriainnaturalenvironments.
Anammoxbacteriacontainauniqueclassoflipids,calledladderanelipidsandherewepresentanovelmethodtodetectviableanammoxbacteriainsedimentsandwastewatertreatmentplantsbasedontheuseofaladderanelipidbiomarker.
Intactladderanephosphatidylcholine(PC)lipidsareanalyzedusingreversed-phaseliquidchromatography–electrosprayionization–massspectrometry.
Followingextractionfromthecomplexsedimentmatrix,reversed-phaseLCisusedtoseparateladderanePClipidsbasedontheirtailgrouphydrophobicityaswellastheiretheroresterlinktotheglycerolbackboneinthesn-2position.
WeinvestigatethepresenceofintactladderanelipidsinnaturalsedimentsdisplayinganammoxactivityandillustratetheuseofaspecificintactmembraneformingPClipidasabiomarkerforviableanammoxbacterialcells.
Thepresentedmethodcanbeusedtoelucidatethewhereaboutsofviableanammoxbacteria,subsequentlyenablinganestimationofanammoxactivity.
Thiswillgreatlyincreasetheknowledgeofanammoxbacteriaandtheirimportanceintheglobalnitrogencycle.
KeywordsLadderanelipids.
LCMS.
Biomarker.
Anammoxbacteria.
NitrogencycleIntroductionItisestimatedthat25%to50%ofthetotalglobalmarinenitrogengas(N2)productionproceedsviatheanaerobicammoniumoxidation(anammox)reaction[1,2].
Locally,theanammoxcontributionmightbeevenhigher[2].
Bacteriaperforminganammoxwerediscoveredinthe1990sinawastewaterreactorandhavesincebeenfoundinamultitudeofenvironments[3–7].
Anammoxbacteriaconvertbiologi-callyavailablenitrogenspecieslikeammonia,nitrateandnitriteintoN2,whichisreturnedtotheatmosphere.
Inthissensetheyarecrucialforsustainingthenitrogenbalanceinourenvironment[8]andgreateffortshavebeenmadeinthesearchforevidenceoftheirpresenceandactivityinnaturalenvironments.
15N-tracerstudieshavebeenusedtodetermineratesofanammoxbymeasuringtheformationof29N2followingincubationof15N-labelednitrate[9,10].
Togetherwithpredictionsfromnutrientandoxygenprofiles,thismethodhasbeenveryusefulinelucidatingwheretheanammoxreactiontakesplace[11–16].
However,studiesusing15N-labelscannotalonebeusedtostatewhethertheanammoxbacteriaareinvolvedornotsincetheanammoxreactionalsocantakeplacethroughalternativeroutes[17].
Using16SrRNAgeneanalysisandmoleculartechniquessuchasfluorescenceinsituhybridization(FISH)andquantitativepolymerasechainreaction(qPCR),anammoxbacteriahavebeenfoundbothinsedimentsandwaterpillars[7,11,13,18,19].
Furthermore,FISHhasbeenasuccessfulmethodologynotonlyfordeterminingthepresencebutalsothediversityofanammoxbacteriaspecies.
However,FISHisamethodthatreliesonfluorescencemeasurementsthusmakingquantitativemeas-urementsproblematicsincemineralandorganicparticlestendtoquenchthefluorescencesignalwhensedimentI.
Lanekoff:R.
Karlsson(*)DepartmentofChemistry,UniversityofGothenburg,Kemivagen10,41296Gothenburg,Swedene-mail:rogerk@chem.
gu.
seAnalBioanalChem(2010)397:3543–3551DOI10.
1007/s00216-010-3913-3samplesareanalyzed.
ThequantitativedeterminationofanammoxbacterialabundancethroughqPCRisalsoassociatedwithissues.
ObservationshaveshownthatbothRNAandDNAcanberelativelystableaftercelllysis[20,21]resultinginenhancedsignalsduetofossilRNAandDNA[22,23].
Hence,thistechniquemightnotreflecttheactualamountofviableanammoxbacteria.
Ourapproachistodeterminethepresenceofanammoxbacteriabyusingaspecificintactmembraneformingphosphatidylcholinelipidfromthefamilyofso-calledladderanelipidsasabiomarkerforthesebacteriainsediment[2,3,6,7,13,24].
Ofallbacterialcellsexaminedtodate,ladderanelipidshaveonlybeenfoundinanammoxbacteria[25].
Ladderanelipidscontainlinearlyconcatenatedcyclo-butanestructuresattheendofthehydrocarbontails,formingaladder-likestructure(Fig.
1).
Thesehighlystrainedstructuresarefoundasphosphatidylcholine(PC),phospha-tidylethanolamine(PE),andphosphatidylglycerol(PG)lipidsaswellasfattyacids(FA)andlysophospholipids[26–28].
Theintactladderanelipidsexistpredominantlywithanetherlinkagetothesn-2positionofaglycerolbackbone,viaanalkylchain,whilethesn-1positionislinkedviaeitheranetheroresterbond[26,29].
Capillarygaschromatography(GC)massspectrometry(MS)hasbeenusedtoanalyzeladderanephospholipidsasladderaneFAMEs(fattyacidmethylesters)afterderivatization[30].
However,thechromatogramsobtainedsufferedfromverybroadpeaks,containingseveralsmallpeakswithalmostidenticalmassspectra.
Thissuggestedthatthestrainedcyclobutanestruc-tureoftheladderaneFAwasthermallyunstableanddegradedduringGCanalysis[30,31].
AnimprovedmethodfortheanalysisofladderaneFAsbasedonnormal-phasehigh-performanceliquidchromatography(NP-HPLC)–atmosphericpressurechemicalionizationtandemmassspectrometry(APCI-MS/MS)waslaterdeveloped[32],aswellasamethodfortheanalysisofaladderanemonoetherPClysolipid[33].
However,asthesetechniquesrelyontheanalysisofFAsandlysophospholipidstheyhavenotprovidedadirectlinktotheamountoflivinganammoxbacteria.
In2009,Jaeschkeetal.
[33]showedthattheC20-[3]-ladderanemonoetherPC,alysophospholipidretain-inganintactPCheadgroupbuthavingonlyonealkylchain,wasabetterbiomarkerforviableanammoxbacteriathantheladderaneFAs.
However,lysophospholipidsaswellasFAsareonlypresentinverylowamountsinlivingcellssincethemajorityofthemembraneisconstitutedofintactlipidscontainingtwonon-polarhydrocarbontailgroupslinkedtoapolarheadgroup.
Duringdegradation,theintactmembraneformingphospholipidsreadilylosetheirheadgroupbutalsooneofthealkylchains,especiallyifitislinkedtotheglycerolbackboneviaanesterbond,resultinginfattyacidsandlysophospholipidsasdegradationproducts.
Inseveralarticles,ithasbeenshownthatintactmembraneformingphospholipidsisareliabletoolfordetectingviablebacteriainsediments[34–40],duetotherapiddecompositionofphospholipidsaftercelldeath.
EarlierworkperformedbyBoumannetal.
[26]hasdemonstratedtheseparationanddetectionofintactladderanelipidsextractedfrombacterialcellculturesandwastewatertreatmentplants,usingnormal-phaseLCandMSdetection.
ThemethodseparatesintactphospholipidsintoPC,PEandPGspecies,butitdoesnotallowforseparationofthelipidsbasedontheirtailgroupsorbondtotheglycerolbackbone.
Earlierstudiesindicatethattherearemanydifferenttypesofladderanelipidspecies,duetoawiderangeofhydrocarbonchainsbondedtotheglycerolbackboneeitherviaanetheroresterbondinthesn-1position.
Whenaspecificintactlipidistobeusedasabiomarkerforviableanammoxbacteria,itisofutmostimportancethatthemethodofuseseparatesthelipidsintoindividualladderanelipidspecies.
Usingnormal-phaseLCseparation,however,itisimpossibletodistinguishbetweenindividualspecieshavingdifferenttailgroupsbutsharingthesameheadgroup.
ThemethodbyBoumannetal.
[26]hasnotbeenusedforanalyzingintactmembraneformingladderanelipidsfromanammoxbacteriainsedimentsamples.
Theanalysisoflipidsoriginatingfromsedimentsiscomplicatedbythepresenceofmanydifferentphospholipidspeciesaswellasamuchhigheramountofcontaminatingorganicmaterialascomparedtowastewatertreatmentsamples.
ThisputshighdemandsontheLCmethodtoseparateoutspecificindividualphospholipidstobeusedasbiomarkersinsedimentsamples.
Here,wehavedevelopedamethodusingaspecificintactmembraneformingladderanePClipid(m/z816,Fig.
1IV)asabiomarkerforviableanammoxbacterialcellsinnaturalsedimentsdisplayinganammoxactivity.
ThemethodenablesPClipidstobeseparatedbasedontheirtailgrouphydrophobicityaswellastheirattachmenttotheglycerolbackbone,whichcanbeeitherthroughanether–etherorester–etherbond.
ThisenablesanalysisofspecificladderanePClipidsinanammoxbacteriafromnaturaloceansediments.
HereweshowthatthepresenceofthespecificladderanePClipidatdifferentdepthsinasedimentcorecorrelateswellwithestablishednutrientandoxygenprofilesinsedimentsaswellasprofilesofanammoxbacterialabundanceestimatedusingmoleculartechniquesincludingFISHandqPCR[33].
ExperimentalsectionChemicalsPhospholipidreferencesamples1,2-didecanoyl-sn-glycero-3-phosphocholine(10PC),1,2-dipalmitoyl-sn-glycero-3-phospho-choline(16PC),1,2-diphytanoyl-sn-glycero-3-phosphocholine(4Me16PC),1,2-di-O-hexadecyl-sn-glycero-3-phosphocholine3544I.
Lanekoff,R.
Karlsson(DE16PC),and1,2-distearoyl-D70-sn-glycero-3-phosphocho-line(D7018PC)werefromAvantiPolarLipids.
ForLCMSmeasurementsLCMSgrademethanol(MeOH)and2-propanol(isopropylalcohol,IPA)werefromRiedel-deHan,formicacidfromFlukaandammoniafromMerck.
Sodium(meta)periodate,chloroform,n-hexane,ammoniumbicarbonate,anddiethyletherwerefromSigma-Aldrich.
Dichloromethane(DCM)anddichlorodimethylsilanewerefromFlukaandNH2solid-phaseextraction(SPE)columns(500mg,3mL)fromStrata.
MaterialsFormethoddevelopment,anammoxbacteriagrowinginbiofilmsonplasticbiofilmcarriers(AnoxkaldnesAB,Lund,Sweden)wereobtainedfromHimmerfjrdsverketswastewatertreatmentplantinStockholm,Sweden.
LadderanePClipidsextractedfromanammoxbacteriabiofilmswereusedasreferencestodeterminetheLCretentiontimeduringsedimentanalysis.
Sedimentcoresamples(dividedintodepths0–3,3–6,6–9,and9–12cm)wereobtainedfromGullmarsfjordenoutsideLysekilonthewestcoastofSwedenandstoredat4°Cforafewdaysuntilanalysiswasperformed.
BiofilmextractionToextractladderanelipids,threedifferentprotocolswereinvestigated.
ThefirstprotocolisdescribedbyZinkandMangelsdorf[41]whereMeOH:DCM:10mMammoniumacetate(AmAc)isusedintheratioof2:1:0.
8.
ThesecondfollowstheFolchprocedureusingamixtureofchloroform:MeOH(2:1)[42]andthethird,fromHaraetal.
[43],usesamixturebasedonn-hexane:isopropanol(3:2).
Priortoextractionthebiofilmsampleswerecentrifugedandcarefullyweighedtoensurethatacomparisonbetweenthedifferentlytreatedbiofilmsamplescouldbemade.
Inallmethodsweusedultrasonication,asdescribedbelow,andextractedthelipidsthreetimeswithacentrifugationstepinbetween.
Supernatantswerecombinedandcollectedinseparateseparationfunnels.
Allbiofilmsamplesweresubjectedtoliquid–liquidextraction(LLE),asdescribedintheoriginalpapers[41–43],andSPE[44].
Thephospholipidfractionswerefullyevaporatedinasilanizedtesttubeunderastreamofnitrogengas,re-dissolvedin200μLmobilephase,andstoredforashortperiodoftimeundernitrogenatmosphereat80°CpriortoLCMSanalysis.
TheFolch[42]andHara[43]procedureswerelessefficientthantheZinkprocedure[41]andwerethusnotusedforthesedimentextractions.
SedimentincubationandextractionAsedimentcorewasobtainedfromGullmarsfjorden(110mbelowseasurface)anddividedintofour3-cm-thicksamplesbetween0and12cmofdepth.
Anaqueoussolutioncontaining5%sodium(meta)periodate(150ml)wasaddedto150gofwetsediment.
Thesedimentslurrywassonicatedfor2min,usinganultrasonicprocessor(SonicsVibracell,Model501)at70%,witha7-spulseanda5-srest.
Theslurrywasincubatedat4°Cforaperiodof1-weektobreakclustersofbacteriaandtolysecells[45,46].
Afterincubation,thesamplewastransferredtotwo250-mLcentrifugeflasks(TeflonFEP,Nalgene)andcentrifugedat3,220gfor15minusinganEppendorfCentrifuge5810R.
Thesupernatantswerecombinedandcollectedinaseparationfunnel.
MeOH(75ml)wasaddedtoeachcentrifugeflaskandthesamplewassonicatedandcentrifugedusingthesameconditionsasabove.
TheMeOHsupernatantswerealsocombinedandcollectedinaseparateseparationfunnel.
Tofurtherextractlipids,theZinkprocedure[41]wasused.
Thesamplewasagaintreatedwithultrasonicationasdescribedabovebutwith75mLofMeOH:DCM:AmAc(2:1:0.
8)threetimesperflaskwithcentrifugationstepsinbetween[41].
Thesupernatantswerecombinedandcollectedinseparateseparationfunnels.
Fig.
1SchematicillustrationsofladderanePClipidsinvestigatedinthisstudy.
(I)Dietherlipidatm/z800,(II)dietherlipidatm/z802,(III)ether/esterlipidatm/z814,(IV)ether/esterlipidatm/z816.
Notethesequentiallylinkedcyclobutane/cyclohexanestructuresattheendofthehydrocarbontailsIntactladderanephospholipidsfromviableanammoxbacteria3545SedimentsamplecleanupprocedurePriortoLLE,DCM,andAmAcwereaddedtoalltheseparationfunnelsuntiltheratiosofthesolventswereMeOH:DCM:AmAc1:1:0.
9.
Afterphaseseparation,theDCMphaseswerecollectedinaroundbottomflask.
ThewaterphasewassubjectedtotwomoreextractionswhereDCM:MeOHwasaddedinaratioof2:1[41].
Again,theDCMphaseswerecollectedandcombinedwiththepreviousextracts.
TheDCMintheroundbottomflaskwasreducedusingarotavaporatorfromBüchi,andfullyevaporatedinasilanizedtesttubeunderaflowofnitrogengas.
Thesamplewasre-dissolvedin12mLof2:1n-hexane:chloroformandstoredundernitrogenatmo-sphereat80°CuntilsubjectedtofurthercleanupbySPE.
FourSPEcolumns(StrataNH2(55μm,70A)500mg,3mL)persamplewereconnectedtoavacuumbox(Resprepvacuummanifoldwith12ports)andactivatedwith12mLMeOH.
Thecolumnswerecondi-tionedwith12mLofn-hexanepriortoloadingthesample.
Awashwith12mLchloroform:IPA(2:1)elutedcholesterylesters,triglycerides,cholesterols,diglycerides,andmonoglycerides.
FAswereelutedwith2%aceticacidindiethylether.
Finally,thephospholipidswereelutedwith12mLofMeOH,combinedandfullyevaporatedinasilanizedtesttubeunderastreamofnitrogengas[44].
Thecleansamplewasre-dissolvedin200μLmobilephase,andstoredundernitrogenatmosphereforashortperiodoftimeat80°CpriortoLCMSanalysis.
LiquidchromatographyandmassspectrometrySeparationwasachievedusingaGeminiC18column(50*2mm,3μmparticlediameter,hybridsilicagelcolumnforreversed-phaseHPLC)withaGeminiC18guardcolumn(4*2mm),fromPhenomenex.
Thetemper-atureofthecolumnwaskeptconstantat30°CusingacolumnovenLCO102fromEcom.
ThemobilephasesinthegradientconsistedofA;4%water,0.
1%formicacid,and0.
1%ammonia(25%)inMeOHandB;isopropanol(IPA)containing0.
1%formicacidand0.
1%ammonia(25%).
Theinitialcompositionwithaflowof0.
4mL/minwas100%Awhichwasloweredto70%Abetween10and20minoftheseparation.
At21min,theflowwasloweredto0.
2mL/minand100%Bwasusedtowashthecolumnfor10minbeforereturningtotheinitialconditions.
Totalanalysistime,includingwashingstep,foreachsamplewas40min.
Theanalyteswereelutedwithin20min.
Analysisofbiofilmsamplesduringmethoddevelopmentwasperformedusinga600SControllerand626PumpfromWatersanda234autosamplerfromGilsoncoupledtoaMicromassQuattroLC,triplequadrupole,withelectrosprayionization(ESI).
MS/MSanalysisenabledparentalionscanningofthePCheadgroup,atm/z184,todetectPClipids.
Argonwasusedascollisiongas,thepotentialoverthecapillarywas4.
0kVandtheconevoltagewas90V.
Sourceblocktemperaturewasheldat150°Canddesolvationtemperatureat300°C.
SedimentsampleanalysiswasperformedusingaPerkin-ElmerSeries200autosamplerandLCpumpcoupledtoaMicromassQToF,quadrupoletime-of-flightwithESIscanningbetweenm/z100and900.
AsaretentiontimereferenceforladderanePClipidsabiofilmsampleextractwasconsecutivelyanalyzedusingtheQToFset-up.
ResultsanddiscussionLadderanePClipidextractionandsamplecleanupAnammoxbacteriahavebeenshowntoformbiofilmswhenculturedinwastewaterreactorsandmostlikelygrowinclustersinoceansedimentsaswell.
Biofilmandclustersaremostlyformedviaaprotein-polysaccharidematrix[47,48]andbiofilmformationmakesitmoredifficulttolysecellsinordertoextractlipidsfromthecellmembranes.
Forthisstudy,severalchemicalandphysicaltreatments,includingboiling,hydrogenperoxideorhypochloratetreatments,mechanicalhomogenizationandsonicationwereattemptedinordertoseparateandlysecellsinbiofilms(datanotshown).
Thesuccessofthesetreatmentsweremonitoredusingfluorescencemicroscopybystainingthecellswith4′,6-diamidino-2-phenylindole.
Furthermore,theextractedladderanePClipidswereanalyzedusingLCMSpostextraction.
Thebestresults,showedbylackofladderanelipidsinsamplesextractedasecondtime,wereachievedwhenanammoxbacteriainbiofilmswereincubatedwitha5%solutionofperiodatepriortoprobesonication,asdescribedintheexperimentalsection[45,46].
NosignificantdifferenceintheladderanePClipidcompositionorquantitydependingontheperiodateincubationtime(2daysversus7days)wereobserved,showingthatnodecompositionoftheladderanePClipidoccurredduringincubation(datanotshown).
InoceansedimentsfromGullmarsfjorden,approximately3%ofallbacteriahavebeenfoundtobeanammoxbacteria[4].
Thelowcellabundanceandthecomplexityofthesedimentsamplematrixputshighdemandsontheextractionmethod.
TooptimizetheyieldofladderanePClipidsintheextractionstep,wecomparedthreeextractionprotocols,usingcarefullyweighedtriplesamplesofanammoxbacteriainbiofilms.
Thesolventmixturesinthecomparedmethodswere;chloroform:MeOH[42],n-hexane:isopropanol[43]andMeOH:DCM:AmAc[41].
Afterextractionandsamplecleanup,allsampleswereanalyzedusingthedescribedLCMSmethod.
InFig.
2,theareaoftheintegratedpeaksof3546I.
Lanekoff,R.
KarlssonthreechosenladderanePClipids,foundatm/z802,814,and816,arecompared.
TheresultshowsthatthehighestyieldofladderanePClipidsisobtainedwhenthemixtureofMeOH:DCM:AmAc[41]isused.
LCmethodforladderanelipidsThedevelopedreversed-phaseLCmethodseparatesPClipidsbasedonthetailgrouphydrophobicity,makingtheladderanelipidselutelast.
Figure3bshowsthechromato-gramgeneratedduringseparationofthefivenon-ladderanePClipidstandardsshowninFig.
3a.
ThisconfirmsthattheLCmethodseparatesthePClipidsbasedonthelengthandhydrophobicityofthehydrocarbontailgroup.
Furthermore,themethodseparatesthePClipidsbasedonthelinktotheglycerolbond,wherethemorepolarether–ester-linkedlipidselutebeforetheether–etherlinkedlipidswiththesametailgroups.
Thisfeatureoftheseparationisespeciallyimportantwhenaspecificladderanelipidisbeingusedasabiomarker.
TheconcentrationsofalllipidsinFig.
3barethesame,buttheresponsevariesconsiderably.
Themostobviousdifferenceisthattheresponseoftheether–etherlipidisseveralmagnitudeslower.
ThisdecreasedsignalintensitycouldbeduetothelowerionizationorfragmentationefficiencyofthedietherPCduringmassspectrometricanalysis[49].
Thehigherdetectionlimitofdietherspeciesencouragedtheuseofanondietherspecieasbiomarker.
InFig.
3c,abiofilmsamplecontaininganammoxbacteriaisanalyzedusingparentalionscanningofthePCheadgroup.
Fig.
3ChromatogramsshowingtheLCseparationofladderanePClipids.
aStructuresoffivenon-ladderanePClipidsusedduringmethoddevelopment.
(1)10PC,(2)16PC,(3)DE16PC,(4)D7018PC,(5)4Me16PC.
bChromatogramofthenon-ladderanePCstandardsatthesameconcentrationanalyzedbyRP-LCQqQusingparentalionscanningofm/z184.
cChromatogramofabiofilmsamplecontaininganammoxbacteriaanalyzedbyRP-LCQqQusingparentalionscanningofm/z184.
Ladderanelipidselutelastinthechromatogram.
Themorepolarether–ester-linkedladderanePClipidsIIIandIVelutebeforetheether–etherlinkedIandIIFig.
2ComparisonofthreeextractionmethodsforladderanePClipidsfoundatm/z802(II),814(III),and816(IV),n=3.
TheRomannumeralsII,III,andIVdenotethestructuresillustratedinFig.
1.
*Denotesasignificantdifference(studentsttest<0.
05)bIntactladderanephospholipidsfromviableanammoxbacteria3547ThealmostidenticalladderanePClipidsatm/z814(III)andm/z816(IV),bothbeingester–etherladderanePClipids,coeluteinthepeakdenotedbyIII,IVinFig.
3c.
Asexpected,theladderanePClipidsatm/z800(I)andm/z802(II),alsoalmostidenticalbutbothdietherladderanePClipids,coelutelaterinthepeakdenotedbyI,II.
ThecoelutionofspeciesIIIandIVaswellasIandIIisacceptablesincetheseanalytesoriginatefromtheanammoxbacteriainconstantrelativeconcentrations,thuskeepinganeventualdiscriminationeffectduringanalysisconstant.
MassspectrometryofladderanelipidsForelectrospraymassspectrometry,PClipidsionizebestinthepositiveionmodeandreadilylosetheirheadgroupatm/z184whensubjectedtocollisioninduceddissociation.
AQqQ,usingparentalionscanningofthem/z184fragmention,wasusedforanalyteconfirmationduringmethoddevelopment.
Figure4ashowsthechromatogramofananammoxbiofilmsample.
ThetwopeakselutinglastcontaintheintactladderanePClipidsI–IVasshowninthecorrespondingmassspectra(Fig.
4b–c).
ThedetectionlimitsoftheladderanePClipidswasfoundtobehigherwhenusingparentalionscanningontheQqQcomparedtoscanningusingtheQToF.
ThiscanpartlybeexplainedbythefactthattheladderanePClipidshavecomparablylonghydrocarbontailgroupswhichmakethemhardtofragment[46].
Therefore,weusedtheQToF,scanningbetweenm/z100and900,todetectladderanePClipidsinsedimentsampleswhichcontainsubstantiallyloweramountsthanbiofilmsamples.
InFig.
4d,asedimentsampleisanalyzedbyRP-LCQToFandtheselectedmasschromatogram,i.
e.
achromatogramwherethem/z816,correspondingtotheladderanePClipidIV,isextractedoutofthetotalionchromatogram,isshown.
ThepeaklabeledasIVistheladderanePClipidatm/z816andthecorrespondingmassspectrumofthatpeakisfoundinFig.
4e.
ThisclearlyshowsthepresenceofanammoxbacteriaintheoceansedimentsamplethatwastakenfromGullmarsfjordenonthewestcoastofSweden.
AscanbeseeninthemassspectrafrombothbiofilmandsedimentsamplestheRP-LCmethodclearlyseparatetheladderanePClipidsfromotherlipidspeciesinthecomplexsedimentsample.
Fig.
4ChromatogramsandmassspectraofintactladderanePClipids.
aChromatogramofabiofilmsamplecontaininganammoxbacteriaanalyzedbyRP-LCQqQusingparentalionscanningofm/z184.
ThetwopeakselutinglateinthechromatogramcontainstheladderanelipidsI–IVasshowninbandc.
bMassspectrumextractedfromthepeakat12.
5mindenotedIII,IVina.
TheintactladderanePClipidsIIIandIVatm/z814and816areidentified.
cMassspectrumextractedfromthepeakat16mindenotedI,IIinA.
TheintactladderanePClipidsIandIIatm/z800and802areidentified.
dSelectedmasschromatogramfortheintactladderanePClipidIVatm/z816inasedimentsampleanalyzedbyRP-LCQToFscanningbetweenm/z100and900.
eMassspectrumextractedfromthepeakat16mindenotedIVind.
TheintactladderanePClipidsIIIandIVatm/z814and816areidentifiedb3548I.
Lanekoff,R.
KarlssonDeterminationofaspecificladderanebiomarkerTheladderanePClipidatm/z816waschosenasabiomarkerforanammoxbacteriabasedontwoimportantcharacteristics.
First;theladderanePClipidatm/z816displaysthehighestabundanceinthemassspectrumwhenusingthedescribedmethod.
Second,theladderanePClipidatm/z816isanether–ester-linkedladderanelipid,whichwehaveobservedtobelessstableinsedimentwhencomparedtotheether–etherlinkedPClipids.
Thelowstabilityinsedimentenvironmentalsosuggeststhatthislipidwouldbettertracklivinganammoxbacteria.
Thiswasinvestigatedbyanalyzingtheamountofester–ether(e.
g.
,m/z816)andether–ether(e.
g.
,m/z800)linkedladderanePClipidsinfreshsedimentandagainafterstoringthatsamesedimentformorethan1monthinarefrigeratorandat20°C.
Theether–esterladderanePClipidswerefoundinreducedlevelsintheoldersedimentscomparedtothefresh,whereastheether–etherladderanePClipidswerefoundinthesameamountsregardlessofsedimenttype.
Thisobservationsuggeststhatthedegradationinsedimentfortheselipidsdoesnotoccuratsimilarratesandthatanester–etherlinkedladderanePClipidisasuitablebiomark-erforlivinganammoxbacteria.
Furthermore,thestabilityoftheether–etherladderanePClipidsoffersanexplanationtowhyotherstudies,analyzingladderanelipiddegradationproducts,havefoundladderaneFAsandladderanelysoli-pidsspreadthroughoutthesedimentdepthprofiles[33].
TodemonstratetheaccuracyoftheladderanePClipidatm/z816asbiomarkerforviableanammoxbacteriawehaveperformedtestsonbothsamplesfromthewastewatertreatmentplantHimmerfjrdsverketinStockholm,SwedenandasedimentcorefromGullmarsfjordenonthewestcoastofSweden.
BiofilmsamplesfromthreeofthereactorsatHimmerfjrdsverketpresumablycontaininganammoxbacteriawereanalyzed.
Thebiomarkerwaspresentinthesamplefromthereactorthatdisplayedanammoxactivity(datanotshown),shownbythegenerationofnitrogengasunderanoxicconditions,indicatingthepresenceofintactlivinganammoxbacteriainthereactor.
DetectingladderanePClipidsinthecomplexmatrixofsedimentisconsiderablymoredifficult.
Thecorrespondingmasschromatogramsofm/z816atthedepths0–3,3–6,6–9,and9–12cmareshowninFig.
5a.
Nutrientandoxygenprofiles,previouslyinvestigatedby,e.
g.
,Jaeschkeetal.
[33],indicatesthatanammoxbacteriaresideintheupperlayerofthesediment,closetotheborderbetweentheanoxicandoxiczone.
Accordingly,theladderanePClipidbiomarkeratm/z816wasfoundatthisdepth,asshowninthedepthprofileinFig.
5b.
Themorestableether–etherladderanePClipidatm/z800was,however,foundatalmosteverydepthinthecore,likelyduetotheslowerdegradationrateforether–etherPClipidsrelativetotheether–esterlipids,asdiscussedabove.
TheamountofladderanePClipidsfoundatthesedepthscanberelatedtoeachothersincetheladderanePClipidshavealinearresponse(integratedpeakareaversusconcentration)intheconcentrationrangeofinterest(datanotshown).
Inthis,examplethedepthsinthecorearerough,andthedepthatwhichtheanammoxbacteriaarefocusedisprobablyalotFig.
5Resultfromthesedimentdepthprofilestudy.
aFouroverlayedmasschromatogramsofm/z816.
Thesedimentsampledepthsareindicatedatthetraces.
ThebottomtraceisareferencesamplefromabiofilmcontainingladderanePClipids.
bDepthprofileofladderanePClipidIVatm/z816(ether–ester),indicatedbythetriangles,andladderanePClipidIatm/z800(ether–ether),indicatedbythecircles.
Depthslicesare3cmthick.
ForladderanePClipidIV,atm/z816,thepeakswerenormalizedtothemaximumareafoundatdepth0–3cmdenoted100%.
ForladderanePClipidI,atm/z800,thepeakswerenormalizedtothemaximumareafoundatdepth6–9cmdenoted100%Intactladderanephospholipidsfromviableanammoxbacteria3549narrower.
ThefindingofanammoxbacteriaatthissiteinGullmarsfjordenisconsistentwithpreviousobservations[4,50].
ConclusionsAmethodhasbeendevelopedtoanalyzeintactladderanePClipidsusingRP-LC-ESI-MS.
Inthisstudy,themethodwasusedtoidentifyaspecificintactladderanePClipidbiomarkerforviableanammoxbacteria.
Thechosenbiomarkerisanether–esterladderanePClipidwhichisfoundatm/z816.
Further,threedifferentmethodsforextractingladderanePClipidshavebeenevaluatedandamultistepsamplecleanupprocedureforladderanelipidsinsedimentsamplesisdescribed.
Whenusingthedescribedmethodonsedimentsamples,wefoundthattheselectedbiomarkerispresentonlyintheupperpartofthesedimentcore.
Thisfindingissupportedbyearlierobservationsthatanammoxbacteriaresidesintheupperpartofthesediment,justbelowtheoxiczone.
Thedescribedmethodprovidesanewtoolfortheanammox-interestedresearchcommunitytostudyviableanammoxbacteriainnaturalenvironments.
Thiswillleadtoanincreasedunderstandingofthepreferredbiogeochemicalenvironmentoftheanammoxbacteriaandhowtheircommunitystructureisaffectedbyongoingenvironmentalchanges.
Furthermore,thelocaliza-tionofviableanammoxbacteriaandaconcomitantobservationofanammoxactivitywillcontributetoagreaterunderstandingoftheglobalnitrogencycle.
AcknowledgmentsTheauthorswouldliketogreatlyacknowledgeProfessorStefanHulthandhisgroupattheDepartmentofChemistry,UniversityofGothenburgforfruitfuldiscussionsaswellashelpwithsamplecollection.
OpenAccessThisarticleisdistributedunderthetermsoftheCreativeCommonsAttributionNoncommercialLicensewhichper-mitsanynoncommercialuse,distribution,andreproductioninanymedium,providedtheoriginalauthor(s)andsourcearecredited.
References1.
DevolAH(2003)Nitrogencycle—solutiontoamarinemystery.
Nature422:575–5762.
KuypersMMM,LavikG,WoebkenD,SchmidM,FuchsBM,AmannR,JorgensenBB,JettenMSM(2005)MassivenitrogenlossfromtheBenguelaupwellingsystemthroughanaerobicammoniumoxidation.
ProcNatlAcadSciUSA102:6478–64833.
JaeschkeA,denCampH,HarhangiH,KlimiukA,HopmansEC,JettenMSM,SchoutenS,DamsteJSS(2009)16SrRNAgeneandlipidbiomarkerevidenceforanaerobicammonium-oxidizingbacteria(anammox)inCaliforniaandNevadahotsprings.
FEMSMicrobiolEcol67:343–3504.
SchmidMC,Risgaard-PetersenN,vandeVossenbergJ,KuypersMMM,LavikG,PetersenJ,HulthS,ThamdrupB,CanfieldD,DalsgaardT,RysgaardS,SejrMK,StrousM,denCampH,JettenMSM(2007)Anaerobicammonium-oxidizingbacteriainmarineenvironments:widespreadoccurrencebutlowdiversity.
EnvironMicrobiol9:1476–14845.
StrousM,FuerstJA,KramerEHM,LogemannS,MuyzerG,vandePas-SchoonenKT,WebbR,KuenenJG,JettenMSM(1999)Missinglithotrophidentifiedasnewplanctomycete.
Nature400:446–4496.
KuypersMMM,SliekersAO,LavikG,SchmidM,JorgensenBB,KuenenJG,DamsteJSS,StrousM,JettenMSM(2003)AnaerobicammoniumoxidationbyanammoxbacteriaintheBlackSea.
Nature422:608–6117.
SchmidMC,MaasB,DapenaA,dePas-SchoonenKV,deVossenbergJV,KartalB,vanNiftrikL,SchmidtI,CirpusI,KuenenJG,WagnerM,DamsteJSS,KuypersM,RevsbechNP,MendezR,JettenMSM,StrousM(2005)Biomarkersforinsitudetectionofanaerobicammonium-oxidizing(anammox)bacteria.
ApplEnvironMicrobiol71:1677–16848.
KuenenJG(2008)Anammoxbacteria:fromdiscoverytoapplication.
NatRevMicrobiol6:320–3269.
ThamdrupB,DalsgaardT(2002)ProductionofN-2throughanaerobicammoniumoxidationcoupledtonitratereductioninmarinesediments.
ApplEnvironMicrobiol68:1312–131810.
WakiM,YasudaT,SuzukiK,SakaiT,SuzukiN,SuzukiR,MatsubaK,YokoyamaH,OginoA,TanakaY,UedaS,TakeuchiM,YamagishiT,SuwaY(2010)Ratedeterminationanddistributionofanammoxactivityinactivatedsludgetreatingswinewastewater.
BioresourceTechnology101:2685–269011.
DalsgaardT,ThamdrupB,CanfieldDE(2005)Anaerobicammoniumoxidation(anammox)inthemarineenvironment.
ResMicrobiol156:457–46412.
DalsgaardT,CanfieldDE,PetersenJ,ThamdrupB,Acuna-GonzalezJ(2003)N-2productionbytheanammoxreactionintheanoxicwatercolumnofGolfoDulce,CostaRica.
Nature422:606–60813.
SchubertCJ,Durisch-KaiserE,WehrliB,ThamdrupB,LamP,KuypersMMM(2006)Anaerobicammoniumoxidationinatropicalfreshwatersystem(LakeTanganyika).
EnvironMicrobiol8:1857–186314.
TrimmerM,NichollsJC,DeflandreB(2003)Anaerobicammo-niumoxidationmeasuredinsedimentsalongtheThamesestuary,UnitedKingdom.
ApplEnvironMicrobiol69:6447–645415.
RysgaardS,GludRN(2004)AnaerobicN-2productioninArcticseaice.
LimnolOceanogr49:86–9416.
RysgaardS,GludRN,Risgaard-PetersenN,DalsgaardT(2004)DenitrificationandanammoxactivityinArcticmarinesediments.
LimnolOceanogr49:1493–150217.
HulthS,AllerRC,GilbertF(1999)Coupledanoxicnitrificationmanganesereductioninmarinesediments.
GeochimCosmochimActa63:49–6618.
RichJJ,DaleOR,SongB,WardBB(2008)Anaerobicammoniumoxidation(Anammox)inChesapeakeBaysediments.
MicrobEcol55:311–32019.
DaleOR,TobiasCR,SongBK(2009)Biogeographicaldistributionofdiverseanaerobicammoniumoxidizing(anammox)bacteriainCapeFearRiverEstuary.
EnvironMicrobiol11:1194–120720.
NannipieriP,CiardiC,BadaluccoL,CasellaS(1986)AmethodtodeterminesoilDNAandRNA.
SoilBiolBiochem18:275–28121.
SchmidM,TwachtmannU,KleinM,StrousM,JuretschkoS,JettenM,MetzgerJW,SchleiferKH,WagnerM(2000)Molecularevidenceforgenusleveldiversityofbacteriacapableofcatalyzinganaerobicammoniumoxidation.
SystApplMicrobiol23:93–10622.
CoolenMJL,OvermannJ(1998)Analysisofsubfossilmolecularremainsofpurplesulfurbacteriainalakesediment.
ApplEnvironMicrobiol64:4513–45213550I.
Lanekoff,R.
Karlsson23.
CoolenMJL,MuyzerG,RijpstraWIC,SchoutenS,VolkmanJK,DamsteJSS(2004)CombinedDNAandlipidanalysesofsedimentsrevealchangesinHolocenehaptophyteanddiatompopulationsinanAntarcticlake.
EarthPlanetSciLett223:225–23924.
ByrneN,StrousM,CrepeauV,KartalB,BirrienJL,SchmidM,LesongeurF,SchoutenS,JaeschkeA,JettenM,PrieurD,GodfroyA(2009)Presenceandactivityofanaerobicammonium-oxidizingbacteriaatdeep-seahydrothermalvents.
ISMEJ3:117–12325.
JettenMSM,vanNiftrikL,StrousM,KartalB,KeltjensJT,OpdenCampHJM(2009)Biochemistryandmolecularbiologyofanammoxbacteria.
CritRevBiochemMolBiol44:65–8426.
BoumannHA,HopmansEC,vandeLeemputI,OpdenCampHJM,vandeVossenbergJ,StrousM,JettenMSM,DamsteJSS,SchoutenS(2006)Ladderanephospholipidsinanammoxbacteriacomprisephosphocholineandphosphoethanolamineheadgroups.
FEMSMicrobiolLett258:297–30427.
RattrayJE,vandeVossenbergJ,HopmansEC,KartalB,vanNiftrikL,RijpstraWIC,StrousM,JettenMSM,SchoutenS,DamsteJSS(2008)Ladderanelipiddistributioninfourgeneraofanammoxbacteria.
ArchMicrobiol190:51–6628.
BoumannHA,LongoML,StroeveP,PoolmanB,HopmansEC,StuartMCA,DamsteJSS,SchoutenS(2009)Biophysicalpropertiesofmembranelipidsofanammoxbacteria:I.
Ladderanephospholipidsformhighlyorganizedfluidmembranes.
BiochimBiophysActaBiomembr1788:1444–145129.
Sinninghe-DamstéJSS,StrousM,RijpstraWIC,HopmansEC,GeenevasenJAJ,vanDuinACT,vanNiftrikLA,JettenMSM(2002)Linearlyconcatenatedcyclobutanelipidsformadensebacterialmembrane.
Nature419:708–71230.
Sinninghe-DamstéJSS,RijpstraWIC,GeenevasenJAJ,StrousM,JettenMSM(2005)Structuralidentificationofladderaneandothermembranelipidsofplanctomycetescapableofanaerobicammoniumoxidation(anammox).
FEBSJ272:4270–428331.
JaeschkeA,LewanMD,HopmansEC,SchoutenS,DamsteJSS(2008)Thermalstabilityofladderanelipidsasdeterminedbyhydrouspyrolysis.
OrgGeochem39:1735–174132.
HopmansEC,KienhuisMVM,RattrayJE,JaeschkeA,SchoutenS,DamsteJSS(2006)Improvedanalysisofladderanelipidsinbiomassandsedimentsusinghigh-performanceliquidchroma-tography/atmosphericpressurechemicalionizationtandemmassspectrometry.
RapidCommunMassSpectrom20:2099–210333.
JaeschkeA,RooksC,TrimmerM,NichollsJC,HopmansEC,SchoutenS,DamsteJSS(2009)Comparisonofladderanephospholipidandcorelipidsasindicatorsforanaerobicammoni-umoxidation(anammox)inmarinesediments.
GeochimCosmo-chimActa73:2077–208834.
AriesE,DoumenqP,ArtaudJ,MolinetJ,BertrandJC(2001)Occurrenceoffattyacidslinkedtonon-phospholipidcompoundsinthepolarfractionofamarinesedimentaryextractfromCarteaucove,France.
OrgGeochem32:193–19735.
SturtHF,SummonsRE,SmithK,ElvertM,HinrichsKU(2004)Intactpolarmembranelipidsinprokaryotesandsedimentsdecipheredbyhigh-performanceliquidchromatography/electro-sprayionizationmultistagemassspectrometry—newbiomarkersforbiogeochemistryandmicrobialecology.
RapidCommunMassSpectrom18:617–62836.
ZinkKG,WilkesH,DiskoU,ElvertM,HorsfieldB(2003)Intactphospholipids—microbial"lifemarkers"inmarinedeepsubsur-facesediments.
OrgGeochem34:755–76937.
MazzellaN,MolinetJ,SyaktiAD,BertrandJC,DoumenqP(2007)Assessmentoftheeffectsofhydrocarboncontaminationonthesedimentarybacterialcommunitiesanddeterminationofthepolarlipidfractionpurity:relevanceofintactphospholipidanalysis.
MarChem103:304–31738.
H.
Ruetters,H.
Sass,H.
Cypionka,J.
Rullkotter,in4thInternationalSymposiumontheInterfacebetweenAnalyticalChemistryandMicrobiology,Bretagne,France,2000,pp.
149–16039.
ZinkKG,MangelsdorfK,GraninaL,HorsfieldB(2008)Estimationofbacterialbiomassinsubsurfacesedimentsbyquantifyingintactmembranephospholipids.
AnalBioanalChem390:885–89640.
RuettersH,SassH,CypionkaH,RullkotterJ(2002)MicrobialcommunitiesinaWaddenSeasedimentcore—cluesfromanalysesofintactglyceridelipids,andreleasedfattyacids.
OrgGeochem33:803–81641.
ZinkKG,MangelsdorfK(2004)EfficientandrapidmethodforextractionofintactphospholipidsfromsedimentscombinedwithmolecularstructureelucidationusingLC-ESI-MS-MSanalysis.
AnalBioanalChem380:798–81242.
FolchJ,LeesM,StanleyGHS(1957)Asimplemethodfortheisolationandpurificationoftotallipidsfromanimaltissues.
JBiolChem226:497–50943.
HaraA,RadinNS(1978)Lipidextractionoftissueswithalow-toxicitysolvent.
AnalBiochem90:420–42644.
KaluznyMA,DuncanLA,MerrittMV,EppsDE(1985)Rapidseparationoflipidclassesinhigh-yieldandpurityusingbondedphasecolumns.
JLipidRes26:135–14045.
HuebnerJ,WangY,KruegerWA,MadoffLC,MartirosianG,BoisotS,GoldmannDA,KasperDL,TzianabosAO,PierGB(1999)IsolationandchemicalcharacterizationofacapsularpolysaccharideantigensharedbyclinicalisolatesofEnterococcusfaecalisandvancomycin-resistantEnterococcusfaecium.
InfectImmun67:1213–121946.
LiuCH,LeeSM,VanLareJM,KasperDL,MazmanianSK(2008)Regulationofsurfacearchitecturebysymbioticbacteriamediateshostcolonization.
ProcNatlAcadSciUSA105:3951–395647.
ElversKT,LeemingK,MooreCP,Lappin-ScottHM(1998)Bacterial-fungalbiofilmsinflowingwaterphoto-processingtanks.
JApplMicrobiol84:607–61848.
O'TooleG,KaplanHB,KolterR(2000)Biofilmformationasmicrobialdevelopment.
AnnuRevMicrobiol54:49–7949.
Al-SaadKA,SiemsWF,HillHH,ZabrouskovV,KnowlesNR(2003)Structuralanalysisofphosphatidylcholinesbypost-sourcedecaymatrix-assistedlaserdesorption/ionizationtime-of-flightmassspectrometry.
JAmSocMassSpectrom14:373–38250.
vandeVossenbergJ,RattrayJE,GeertsW,KartalB,vanNiftrikL,vanDonselaarEG,DamsteJSS,StrousM,JettenMSM(2008)Enrichmentandcharacterizationofmarineanammoxbacteriaassociatedwithglobalnitrogengasproduction.
EnvironMicrobiol10:3120–3129Intactladderanephospholipidsfromviableanammoxbacteria3551
- Priorqqq258.com相关文档
- dioxinsqqq258.com
- Intramolecularqqq258.com
- 发行人qqq258.com
- 前言本目录对外公开发布,作为全年中心城市用人单位招引人才及兑现补贴的依据.
- 湖南qqq258.com
- throughputqqq258.com
Contabo美国独立日促销,独立服7月€3.99/月
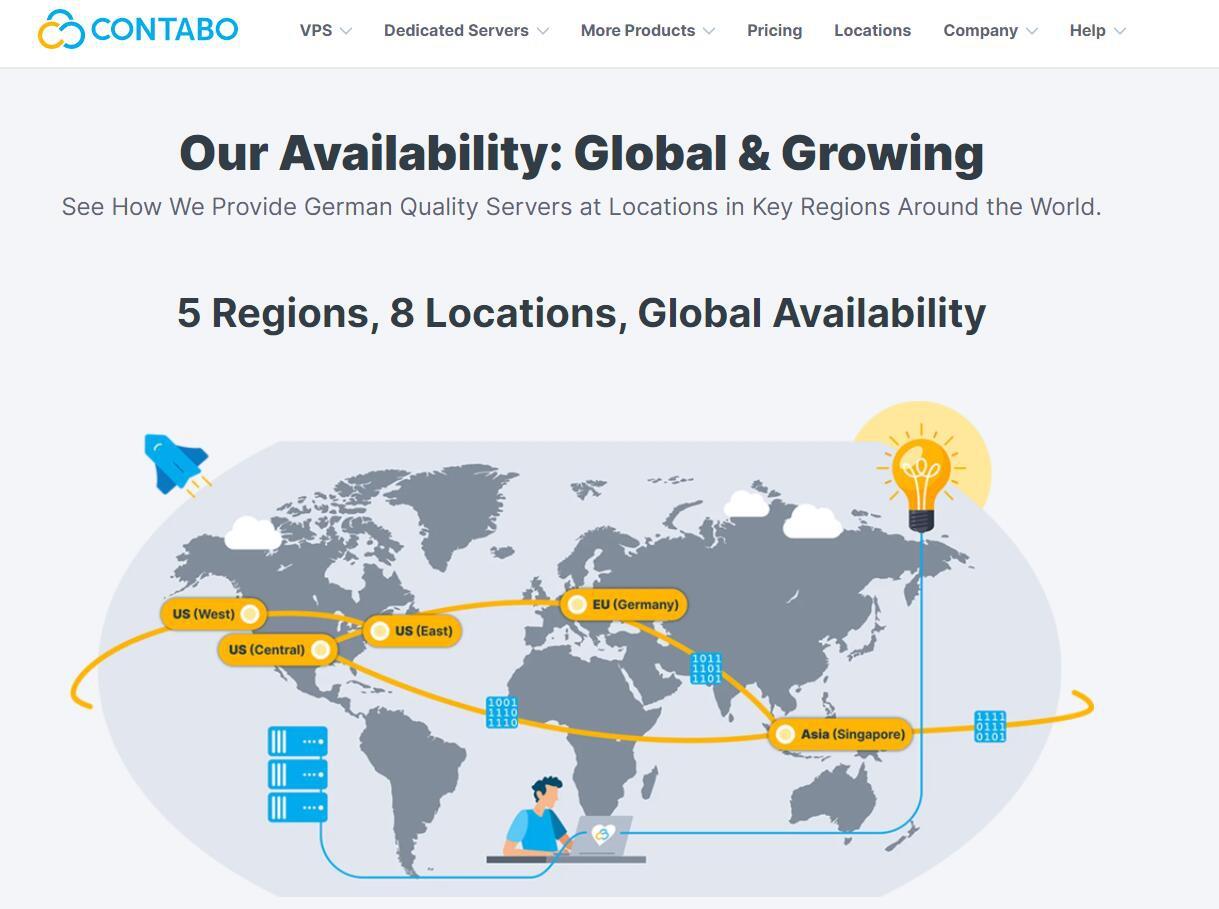
Contabo自4月份在新加坡增设数据中心以后,这才短短的过去不到3个月,现在同时新增了美国纽约和西雅图数据中心。可见Contabo加速了全球布局,目前可选的数据中心包括:德国本土、美国东部(纽约)、美国西部(西雅图)、美国中部(圣路易斯)和亚洲的新加坡数据中心。为了庆祝美国独立日和新增数据中心,自7月4日开始,购买美国地区的VPS、VDS和独立服务器均免设置费。Contabo是德国的老牌服务商,...
georgedatacenter39美元/月$20/年/洛杉矶独立服务器美国VPS/可选洛杉矶/芝加哥/纽约/达拉斯机房/
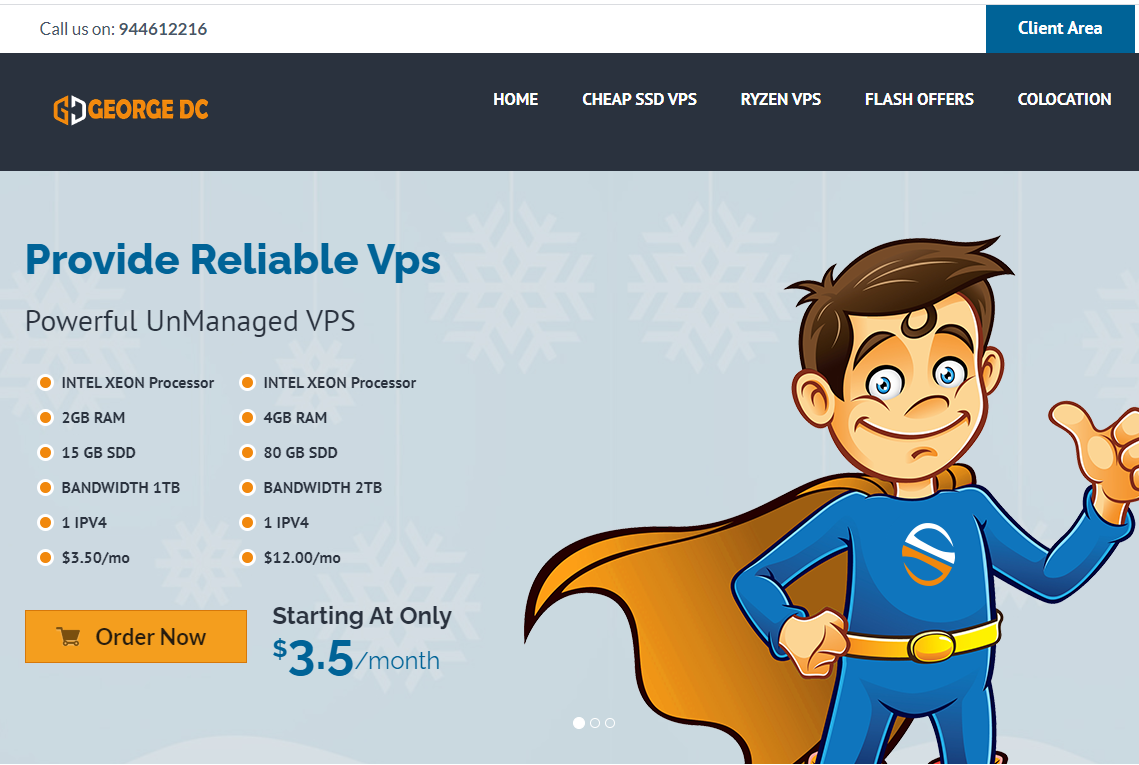
georgedatacenter这次其实是两个促销,一是促销一款特价洛杉矶E3-1220 V5独服,性价比其实最高;另外还促销三款特价vps,georgedatacenter是一家成立于2019年的美国VPS商家,主营美国洛杉矶、芝加哥、达拉斯、新泽西、西雅图机房的VPS、邮件服务器和托管独立服务器业务。georgedatacenter的VPS采用KVM和VMware虚拟化,可以选择windows...
妮妮云(119元/季)日本CN2 2核2G 30M 119元/季
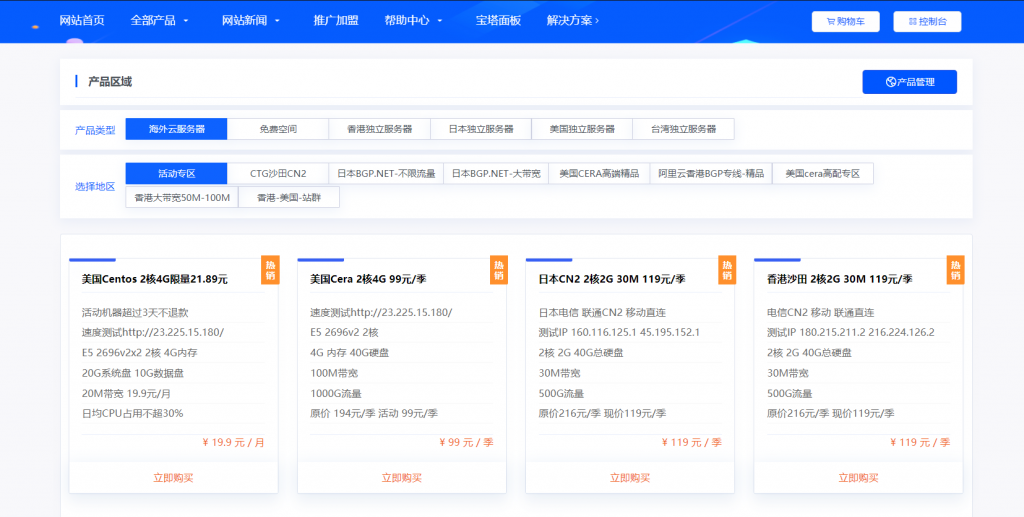
妮妮云的知名度应该也不用多介绍了,妮妮云旗下的云产品提供商,相比起他家其他的产品,云产品还是非常良心的,经常出了一些优惠活动,前段时间的八折活动推出了很多优质产品,近期商家秒杀活动又上线了,秒杀产品比较全面,除了ECS和轻量云,还有一些免费空间、增值代购、云数据库等,如果你是刚入行安稳做站的朋友,可以先入手一个119/元季付的ECS来起步,非常稳定。官网地址:www.niniyun.com活动专区...
qqq258.com为你推荐
-
微盟赔付方案如果合同没有约定违约金赔偿方案,违约怎么索赔·云计算什么叫做“云计算”?嘉兴商标注册个人如何申请商标注册老虎数码虎打个数字甲骨文不满赔偿如果合同期不满被单位辞退,用人单位是否需要赔偿原代码求数字代码大全?psbc.com怎样登录wap.psbc.com百花百游百花净斑方多少钱一盒同一ip网站最近我们网站老是出现同一个IP无数次的进我们网站,而且是在同一时刻,是不是被人刷了?为什么呀?www.299pp.com免费PP电影哪个网站可以看啊