0.21网易yeah
网易yeah 时间:2021-02-16 阅读:()
Limnology(2019)20:163–171https://doi.
org/10.
1007/s10201-018-0562-2RESEARCHPAPERCharacterizationofphosphorusinalgaefromaeutrophiclakebysolution31PnuclearmagneticresonancespectroscopyWeiyingFeng1·CuicuiLi1,2,3·ChenZhang1·ShashaLiu1·FanhaoSong1·WenjingGuo1·ZhongqiHe4·TingtingLi1·HaiyanChen1Received:4July2018/Accepted:12September2018/Publishedonline:4October2018TheAuthor(s)2018AbstractTheidentificationandquantificationofphosphorus(P)compoundsderivedfromalgalbiomassarecrucialforabetterunderstandingofalgalPdynamicsinlakeecosystems.
QuantityandspeciesofPinalgaecollectedfromChaoLake(atypi-calultra-eutrophiclake)inChinawereanalyzedbychemicalanalysisand31PNMR.
TotalP(TP)inalgaebiomassrangedfrom2671to5385mgkg1ofdrymatter.
ProportionoforganicP(Po)accountedfor78.
3±2.
6%inalgaebiomasscollectedfromthewesternpartofChaoLake,whichwashigherthanthat(64.
7±1.
4%)intheeasternpartofthelake.
EightPspeciesincludinginorganicPspecies(orthophosphateandpyrophosphate)andPospecies(fivemonoestersPanddiestersP)wereidentifiedinNaOH–EDTAextractsofalgalsamples.
MonoestersPaccountedfor48.
4%inextractedTP,whichwasthemaincomponentofPo.
β-glycerophosphateswerethelargestcomponentofmonoestersP,whichaccountedfor22.
6%inextractedTP.
ThisstudyimprovedknowledgeonthemechanismofthecyclingofendogenousPintheaquaticsystemandwouldbehelpfulindevelopingastrategyforcontrolofrepeatedalgaebloomsineutrophicChaoLake.
Keywords31PNMR·Algae·Organicphosphorus·Lake·EutrophicationIntroductionAlgaebloomsareseriousenvironmentalproblemsaroundtheworld,especiallyindevelopingcountries(Kagalouaetal.
2008;Pernet-Coundrieretal.
2012).
Algaebloomsoccurinanaquaticenvironmentiftoomuchphosphorus(P)entersthesystem(Gilesetal.
2015).
Foraeutrophiclake,releaseofPfromdeadalgaeisanimportantnutrientsourcethatwillsupportcontinuousalgalbloomsinlakes(Lietal.
2009;Fengetal.
2016a).
Algaearenotusuallycollectedbyhumansbecausetheyarenotvaluable,sothatthedebrisisallowedtodecomposeinsitu.
Thedecomposi-tionofalgalresiduesaffectsthebio-cyclingandreleaseofP,increasingtheriskofresurgenceofalgalblooms(Fengetal.
2016b;Lehmanetal.
2017).
DecayingalgaldebrisreleasesbothinorganicP(Pi)andPo.
ThePoconstituentsneedtobehydrolyzedtobioavailablePobyvariousenzymes(Fengetal.
2016b).
Therefore,theformsandconcentrationsofPinalgaeshallbeevaluatedwhilealgaedebrisisdecomposing.
However,untilnow,studiesofthespecies,concentrations,andeffectsofPoinalgaeofeutrophiclakeshavebeenlim-itedbecauseofthecomplexityandlimitationsofanalyticalmethods(Turneretal.
2005;Belletal.
2017).
Typicalanalyticalapproaches,suchasenzymaticreac-tivity,highperformanceliquidchromatography(HPLC),andmassspectrometryarebasedonoperationaldefini-tionssothattheycannotdiscernPclassesatamolecularlevel(Suzumura2005;Baldwin2013;Karl2014).
Phos-phorus-31nuclearmagneticresonancespectroscopy(31PHandlingEditor:RichardSheibley.
WeiyingFengandCuicuiLiareco-leadauthors.
*ChenZhangzhangchen0353@yeah.
net*HaiyanChenchenhy@craes.
org.
cn1StateKeyLaboratoryofEnvironmentalCriteriaandRiskAssessment,ChineseResearchAcademyofEnvironmentalSciences,Beijing100012,China2GuangzhouInstituteofGeochemistry,ChineseAcademyofSciences,Guangzhou510640,China3UniversityofChineseAcademyofSciences,Beijing100049,China4USDAARS,SouthernRegionalResearchCenter,NewOrleans,LA70124,USA164Limnology(2019)20:163–171NMR)isanon-destructive,non-invasivetechniqueforiden-tifyingchemicalformsinvariousenvironmentalsamples.
Solid-state31PNMRisfocusedoninorganicPcompounds,solution31PNMRismainlyusedtodetermineorganicPcompounds(Turneretal.
2005;Heetal.
2011;Abdietal.
2014;Srensenetal.
2014).
SeveralPcompoundshavebeendetectedbysolution31PNMR,includingphosphonates,orthophosphate,monoestersP,diestersP,pyrophosphatesandpolyphosphates(Heetal.
2007;Turneretal.
2012;Zhuetal.
2013).
Generally,monoestersPrepresentawiderangeofimportantPocompounds,suchasinositolphos-phateandsugarphosphates(Heetal.
2007;Dooletteetal.
2009;Jaroschetal.
2015).
Therefore,itisanidealtechniqueforanalyzingPospeciesinalgaeofeutrophiclakes,asitwouldnotonlyprovideimportantinformationpertainingtoPbiogeochemicalcyclinginlakeecosystems,butalsoyieldabundantinsightintoidentitiesofspecificPcompounds.
ChaoLake(31°25′28″–31°43′28″N,117°16′54″–117°51′46″E)isoneofthefivelargestfreshwa-terlakesinChina.
ItissituatedonthefloodplainsbetweentheYangtzeRiverandHuaiRiverinthecentralAnhuiProv-inceofeasternChina(Zanetal.
2010;Tangetal.
2015).
ChaoLakeisatypicalshallowlakewithameandepthof3mandasurfaceareaof780km2anddrainageareaof13,350km2(Wangetal.
2013).
Duetotherapidincreaseinanthropogenicactivitiesinthelake'swatershedoverrecentdecades,thelakehassufferedfromseriouspollution,eutrophicationandalgaeblooms(Xuetal.
2005).
Asamat-teroffact,sincethemid1980s,algaebloomshaveoccurredeachyearinChaoLake(ChenandLiu2014).
Inordertoassesstheenvironmentalrisksofalgaeontheeutrophiclake,thisstudyanalyzedthePspeciesinalgaebysolution31PNMR,andbasedontheknowledgeobtained,predictedthePbioavailabilityofthealgalbiomassinaquaticenvironments.
MaterialsandmethodsStudysitesandsamplecollectionSamplesofalgaewerecollectedfromsixsitesinChaoLakeinSeptember2015(Fig.
1).
Thesesamplingsiteswerelocatedindifferenteutrophicareas.
Generally,ChaoLakeisdividedintotwoparts:thewesternpart(samplesC1andC2)andtheeasternpart(samplesC3,C4,C5andC6)alongthelineofZhongmiao–Mushan–Qitouzui,asshowninFig.
1.
Thequalityofwaterwasworstinthewesternpartofthelakeandgraduallybecamebetterfromwesttoeast(Zhuetal.
Fig.
1MajorinletsandoutletsofChaoLakewithalgalsamplingsites165Limnology(2019)20:163–1712006;Tangetal.
2015).
Thelake'sannualmeanconcentra-tionsoftotalnitrogen(TN)andTPapproached2.
85and0.
26mgl1,respectively,andtheannualmeanchlorophyll-areachedupto25.
6μgl1(Lietal.
2015).
Algaewerecol-lectedbyuseofaplanktoncollector(HB403-BWS).
Sam-pleswereplacedinsealedbagsandputiniceboxesimme-diately.
Thesealgalsampleswerefreeze-dried,ground,thenpassedthrougha2-mmsievebeforebeingstoredat20°C(Fengetal.
2016a).
ThedominantspeciesinChaoLakewasMicrocysticaeruginosawithanappearancefrequencyof90.
9%(Yangetal.
2011).
ExtractionofPandchemicalanalysisSamplesofalgaewereextractedbyuseofopti-mizedNaOH–EDTAextractingagent(mixturesof0.
5moll1NaOHand25mmoll1EDTA)witharatioof150:1(mlg1),andthemixtureswereshakenfor18hatroomtemperature(Cade-MenunandPreston1996;Fengetal.
2016b).
Theextractingsolutionswerecentrifuged(8000*g)for30min,andfilteredthrough0.
45-μmglass-fiberfilters(WhatmanGF/C).
Extractabletotalphosphorus(ETP)afterdigestionandfreemolybdatereactivephos-phorus(MRP)weremeasuredusingthemolybdenumbluemethod(HeandHoneycutt2005).
Extractableorganicphos-phorus(EOP)wascalculatedbythedifferencebetweenETPandMRP.
Theremainingextractswerefreeze-driedforsolu-tion31PNRMspectroscopyanalysis.
Percentagesofcarbon(C)andnitrogen(N)inalgaeweredeterminedbyuseofanelementalanalyzer(ElementarvariomacroEL,Germany).
Totalphosphorus(TP)andinorganicphosphorus(Pi)weredeterminedbytheSMTmethoddescribedbyRubanetal.
(2001).
Organicphosphorus(Po)inalgaesampleswascalculatedbythedifferencebetweenTPandPi.
TherewerethreereplicationsforextractionofPandchemicalanalysis.
Datawerecheckedfordeviationsfromnormalityandhomogeneityofvariancebeforeperformingstatisticalanalyses.
31PNMRanalysisA100-mgsampleoffreeze-driedalgalextractswasground,andthenredissolvedin1ml1moll1NaOH+0.
1moll1EDTAand0.
2mlD2O.
Afterultrasonicationfor30minandequilibrationfor5min,2%(v/v)ofbicarbonatebuffereddithionite(0.
11moll1NaHCO3+0.
11moll1Na2S2O4)wasaddedtotheextracttoreduceinterferencefrompara-magneticions,suchasFeandMn(Heetal.
2009;GilesandCade-Menun2014).
ThepHofthesupernatantsolutionwasadjustedusing10moll1NaOHtoensureapH>12.
Thesupernatantsolutionwascentrifuged(8000*g)for30minandtransferredtoa5-mmNMRtube.
Solution31PNMRspectrawereacquiredat24°ConaBrukerAVANCE400MHzspectrometerata31Pfrequencyof161.
98MHz,usinga90°pulse,a5srelaxationdelayanda0.
21sacqui-sitiontime,similartotheparametersusedinFengetal.
(2016b).
Thescantimeforeachsamplewasmorethan15h.
PeakareaswerecalculatedbyintegrationandcompletedusingMestrelabMNovav.
10.
SpikingexperimentsThepeakofspecificmonoestersPforms(i.
e.
,glu-cose6-phosphate,riboncleotides,α-glycerophosphate,β-glycerophosphate,myo-inositolhexaphosphate)needstobeconfirmedwithspikingexperiments(Fig.
4).
MethodsforidentifyingspecificPformsinNMRspectraofsoilandotherenvironmentalsamplesarewell-establishedandhavebeenusedformanyyears(SmernikandDougherty2007;Heetal.
2011;McDowellandHill2015),combinedwithPcom-poundlibrariesdevelopedbyTurneretal.
(2003);Dooletteetal.
(2009)andCade-Menun(2015).
StandardsamplesofmonoestersPwerepurchasedfromSigma-Aldrich.
Spikedsampleswereanalyzedby31PNMRasdescribedabove.
MonoestersPcompoundswereidentifiedbytheirchemicalshifts,withtheorthophosphatepeakinallspectrastand-ardizedto6.
0ppm.
SpectralprocessingwasdoneusingMestReNovesoftwareversion9.
0.
1(MestReNoveResearchSL).
ResultsandanalysisNutrients(C,NandP)indebrisofalgaeCompositionofC,NandtheirratiosindebrisofalgaeareshowninTable1.
PercentagesofCrangedfrom31.
5to50.
3%withameanvalueof41.
9%inthealgaesamplesfromChaoLake.
ContentofNwas5.
2–7.
8%withameanvalueof6.
4%.
BothcontentsofCandNofalgaewasgreaterthanthoseofaquaticmacrophytes,whichhasbeenwidelyreportedpreviously(Zhongetal.
2012;Quetal.
2013,Fengetal.
2016a).
TheratioofC:Nwasagoodpredictorofdeg-radationinalgaeandaquaticmacrophyteswithalowerratioofC:Nformaterialmorereadilydegradation(Reitzeletal.
2006).
Inthisstudy,theratiosofC:Ninalgaefromthewest-ernlake(7.
1±0.
3)werehigherthanthoseoftheeasternlake(6.
4±0.
3),suggestinghigherlabilityofthealgaldebrisoftheeasternlake.
TheratioofC:Nofaquaticmacrophytes(average12.
9)(Fengetal.
2016a)wasgreaterthanthatofalgaeinChaoLake(average6.
7)(Table1).
Therefore,thealgaedecomposedmoreeasilythanaquaticmacrophytesinthesamelake.
Thiswasalsoconsistentwiththeresultsofpreviousstudies(Liuetal.
2016).
ContentsofTPindebrisofalgaerangedfrom2671to5385mgkg1drymass(dm)withameanvalueof166Limnology(2019)20:163–1713936mgkg1.
ThegreatestconcentrationofTPindebrisofalgaewasobservedinsampleC6.
Thismaybebecauseoftwomajorinflowingrivers(e.
g.
,theZhegaoriverandShuangqiaoriver)withheavynutritioninputs(Tangetal.
2015).
ContentsofTPinthesurfacesedimentsfromChaoLakeonlyrangedfrom420to1090mgkg1withameanvalueof687mgkg1(ZhangandXing2013).
ContentofTPindebrisofalgaefromChaoLakewasapproximately5timeshigherthanTPinsurfacesediments.
Therefore,deadalgae-derivedPmightbeanimportantsourceofbioavail-ablePforrepeatedalgalbloomingineutrophiclakessuchasChaoLake.
ConcentrationofPointhesealgalsamplesrangedfrom2107to4136mgkg1withameanvalueof2783mgkg1.
ThegreatestconcentrationofPowasalsoobservedintheheavilypollutedregion(i.
e.
,sampleC6).
TheproportionofPoinalgaeofthewesternlake(78.
3%±2.
6%)washigherthanthatintheeasternlake(64.
7%±1.
4%).
ThemeanratioofPo/TPofthesixalgalsampleswas69.
2%inChaoLake(Table1).
ThisvaluewasgreaterthanthatofalgaefromTaiLake(meanratioofPo/TP57.
8%)(Fengetal.
2016b).
ThepreviousstudiesreportedthatPocouldbecon-vertedtobioavailableP(e.
g.
,HPO42)foralgaebloomingthroughaseriesofredox-drivensolubilizationreactionsandphosphatase-mediatedhydrolyticprocesses(WangandPant2010;Zhuetal.
2015).
Thus,webelievedthatPoindebrisofalgaefromChaoLakepossessedlargerbioavailabilitypotentialthanalgaefromTaiLakepertheirdifferenceinPo/TPratios.
NaOH–EDTAextractablePfromalgaeInthisstudy,contentsofNaOH–EDTAextractableTPrangedfrom1292to2774mgkg1,withanextractionefficiencyof40.
5–78.
3%,andcontentsofextractablePofromalgaerangedfrom351to1141mgkg1,withanextrac-tionefficiencyof13.
8–54.
2%(Table1).
TherecoveriesofPoindebrisofalgaefromChaoLakewerelowerthanthosewithpurealgaesuchasMicrocystis,Chlorellavulgaris,andSprilinaplatensis(Fengetal.
2016a).
However,theextrac-tionefficienciesofPoofalgaefromChaoLakeweresimilartothoseofparticulatePfromTaiLake(23–56%)(Baietal.
2017)andofsedimentsfromHaiheRiver(30–73%)(Zhangetal.
2017).
TheextractionefficiencyofTPfromalgaefromChaoLakewassimilartothatofsoilsandsediments(49–83%)(Xuetal.
2005;Tangetal.
2015).
Multiple-stepextractions(e.
g.
,additionalorsequentialHClextraction)(Heetal.
2008;Cade-Menun2015;Zhuetal.
2016;Liuetal.
2017)seemneededtoincreasethePrecoveryfromthesealgalsamples.
Solution31PNMRspectraofNaOH–EDTAextractsofalgaeEightmainPspeciesincludinginorganicPspecies(orthophosphateandpyrophosphate)andPospecies(fivemonoestersPanddiestersP)wereidentifiedintheNaOH–EDTAextractsofthesixalgalsamplesbysolution31PNMR(Fig.
2;Table2).
Thepeakoforthophosphatewasat6.
00ppminthe31PNMRspectra,monoestersPwasat3.
33–5.
49ppm,diestersPwasat0.
69to0.
31ppm,andpyrophosphatewasat4.
21to4.
12ppm(Fig.
2).
ThesumoforthophosphateandmonoestersPinETPaccountedformorethan93%ofETP(Table1;Fig.
3b).
Withthepeakoforthophosphateasthelargestsignalinthese31PNMRspectra.
Thecontentoforthophosphatewasbetween615.
1and1331.
7mgkg1,andaccountedTable1ContentsofC,N,andPinalgaeandtheirNaOH–EDTAextractionefficiencyinChaoLakeETPextractabletotalphosphorus,EOPextractableorganicphosphorusaMean±standarddeviation(n=3)bValuesinbracketsshowthepercentageofETP,EOPinNaOH–EDTAextractstoTPandPointheunextractedalgaepowders,respectivelySamplesCoordinatesOriginalalgaepowdersNaOH–EDTAextractalgaesamplesC(%)N(%)C:NTP(mgkg1)Po(mgkg1)Po/TP(%)ETP(mgkg1)EOP(mgkg1)C131°38′3.
52″E,117°21′16.
74″N44.
8±2.
5a6.
7±0.
56.
7±0.
34173±5163372±12680.
8±1.
61864±126(44.
7)b785±124(23.
3)bC231°34′3.
50″E,117°24′51.
80″N39.
5±0.
65.
3±0.
87.
5±0.
24059±1253072±43275.
7±3.
51757±214(43.
3)464±52(15.
1)C331°30′9.
64″E,117°28′56.
52″N38.
3±1.
85.
4±0.
27.
1±0.
43787±2562536±5967.
0±2.
51572±56(41.
5)351±98(13.
8)C431°27′56.
84″E,117°34′50.
00″N31.
5±3.
55.
2±0.
16.
1±0.
12671±1981476±12355.
3±1.
41292±89(48.
4)567±15(38.
4)C531°33′27.
70″E,117°36′46.
18″N50.
3±2.
97.
8±0.
86.
5±0.
43543±1122107±7859.
5±0.
92774±21(78.
3)1141±215(54.
2)C631°36′10.
25″E,117°47′38.
55″N46.
9±4.
67.
8±0.
76.
0±0.
25385±2904136±1976.
8±0.
82183±51(40.
5)1066±164(25.
8)167Limnology(2019)20:163–171for41.
5–54.
0%ofETP(Table1;Fig.
3).
PolyphosphateswerenotdetectedinanyalgaeofChaoLake.
MonoestersPcomprisedthelargestPofractionwithNaOH–EDTAextracts,andaccountedfor48.
4%(average)inETP(Table2;Fig.
3b).
Throughthespikingexperi-ments(Fig.
4),thepeakat4.
88±0.
02ppmwasassignedtoα-glycerophosphatesandthepeakat4.
50±0.
02ppmwasassignedtoβ-glycerophosphates,basedonTurnerandRichardson(2004)andDooletteetal.
(2009);thepercentagesofα-andβ-glycerophosphatesinETPwere3.
4%(65.
5mgkg1)and22.
6%(432.
7mgkg1),respec-tively.
β-glycerophosphateswerethelargestcomponentofmonoestersP.
Thepeakat4.
32±0.
01ppmwasassignedtoribonucleotides(Heetal.
2011)andthepercentageofribonucleotidesinETPwas10.
6%(218.
2mgkg1).
Thepeakat5.
12±0.
01ppmwasassignedtoglucose6-phos-phate(Cade-Menun2015),thepercentageofwhichinETPwas1.
9%(38.
7mgkg1).
Inaddition,afewofthepeaksinthemonoestersPregionwereunidentified,becausechemicalshiftswerestronglyinfluencedbysubtlediffer-encesamongsamplesforviscosity,pH,saltsandparamag-neticions(Youngetal.
2013;Abdietal.
2014;Heetal.
2011;Gilesetal.
2015).
UnidentifiedmonoestersParedefinedas'othermonoestersP'inthisstudy.
TheseothermonoestersPaccountedfor9.
9%ofETPinalgaeofChaoLake(Table2).
PyrophosphatewasdetectedinmostsamplesexceptthesampleC1,whichisconsistentwithanumberofotherstud-ies(Bedrocketal.
1995;Mahieuetal.
2000;Heetal.
2011).
Inotherliterature,polyphosphatesandpyrophosphatewere6420-2-4C2C1OrthophosphatePyrophosphateMonoestersPC3^C5C4^C6DiestersPChemicalshift(ppm)Fig.
2Solution31PNMRspectraofNaOH–EDTAextractsofalgaeinChaoLakeTable2ConcentrationsofPcompoundsinNaOH–EDTAextractsofthealgaedeterminedbysolution31PNMRPiinorganicP,PoorganicP,TotalPothesumofmonoestersPanddiestersP,n.
dnotdetectedaValuesinparenthesesarepercentagesofindividualPcompoundsinNaOH–EDTAextractsTPAlgaePi(mgkg1)Po(mgkg1)OrthophosphatePyrophosphateMonoestersPDiestersPTotalPoGlucose6-phos-phateRibonucleotidesα-glycerophosphateβ-glycerophosphateOthermonoestersPTotalmonoestersPC1818.
2(43.
9)an.
d32.
7(1.
8)204.
6(11.
0)60.
6(3.
2)526.
9(28.
3)220.
9(11.
9)1045.
7(56.
1)n.
d1045.
7(56.
1)C2938.
1(53.
4)115.
4(6.
6)42.
2(2.
4)144.
5(8.
2)63.
8(3.
6)322.
7(18.
4)130.
4(7.
4)703.
5(40.
0)n.
d703.
5(40.
0)C3849.
1(54.
0)15.
8(1.
0)n.
d137.
6(8.
7)49.
2(3.
1)325.
2(20.
5)201.
5(12.
7)714.
1(45.
0)n.
d714.
1(45.
0)C4615.
1(47.
6)65.
8(5.
1)15.
4(1.
2)131.
2(8.
8)44.
9(3.
5)297.
7(23.
0)131.
0(10.
1)604.
4(46.
6)9.
2(0.
7)611.
4(47.
3)C51331.
7(48.
0)87.
9(3.
2)49.
3(1.
8)487.
4(17.
6)103.
9(3.
7)636.
6(22.
9)67.
9(2.
4)1346.
1(48.
5)9.
3(0.
3)1354.
4(48.
8)C6905.
6(41.
5)32.
6(1.
5)92.
4(4.
2)203.
8(9.
3)70.
6(3.
2)487.
2(22.
3)329.
6(15.
1)1184.
0(54.
2)61.
6(2.
8)1245.
2(57.
0)168Limnology(2019)20:163–171detectedinsomesamples,butnotnecessarily(Busatoetal.
2005;Fengetal.
2016b).
DiscussionIdentificationofinositolhexaphosphate(IHP)inalgaeofChaoLakeTheinositolhexaphosphate(IHP)stereoisomers(scyllo-,myo-,chiro-,neo-IHP)wereimportantmonoestersPcom-ponentsinmanyenvironmentalsamples(Turneretal.
2012;Cade-Menun2015).
Eachofthesecompoundscontainssixphosphates,andtheconformationofthosephosphategroupscausesthemtohavemultiplepeaksinasinglespectrum,inanarrangementspecifictoeachcom-pound.
Theonlyexceptionisscyllo-IHP,whichhasonepeakforthesixphosphates;thepeakat3.
55±0.
02ppmwasassignedtoscyllo-IHP,basedonTurnerandRichard-son(2004)andDooletteetal.
(2009).
NoneofthespectrashowninFig.
2haveapeakat3.
55ppm,sothescyllo-IHPwasnotpresentinalgaeofChaoLake.
Myo-IHPhasfourpeaksina1:2:2:1arrangement(withrespecttopeakareas),withpeaksat5.
79±0.
01,4.
88±0.
01,4.
55±0.
01and4.
42±0.
01ppm(inFig.
4e).
ThreeofthesepeakswerenotpresentinalgaesamplesinChaoLake(Fig.
2),somyo-IHPwasnotpresentinalgaeofChaoLake.
Thefactthatmyo-IHPhadnotbeenobservedinalgaeinapreviousstudyisconsistentwiththeobservationofthisstudy(Fengetal.
2016a).
Forchiro-IHP[ineitherthe4equatorial/2axial(4e/2a)conformationorthe2e/4aconformation],threepeaksmustbeclearlyvisible,ina2:2:2arrange-ment,andthediagnosticpeaksforeachareat6.
2–6.
5ppm(Cade-Menun2015);however,noneofthespectrashowninFig.
2haveanypeaksbetween6.
0and7.
0ppm,sothesecompoundswerenotpresentinalgaeofChaoLake.
Inaddition,neo-IHPrequirestwopeakstobepresent,ina4:2arrangement,sothatthepeakat6.
4±0.
01ppmistwiceaslargeastheoneat4.
3±0.
01ppm.
Giventhattherewerenopeaksbetween6.
0and7.
0ppm,itcanbe050010001500200025003000C1C2C3C4C5C6&RQFHQWRIPDMRU3W\SHVPJ3NJ$OJDOVDPSOHV(a)(b)(c)(d)pyrophosphatediestersPmonoestersPorthophosphate0%20%40%60%80%100%C1C2C3C4C5C63HUFHQWDJHRIPDMRU3W\SHVLQ(73$OJDOVDPSOHVorthophosphatemonoestersPdiestersPpyrophosphate0200400600800100012001400C1C2C3C4C5C6&RQFHQWRIPRQRHVWHUV3$OJDOVDPSOHVOthermonoestersPβ-glycerophosphateα-glycerophosphateribonucleotidesglucose6-phosphate0%20%40%60%80%100%C1C2C3C4C5C63HUFHQWDJHRIPRQRHVWHUV3LQ(73$OJDOVDPSOHVOthermonoestersPβ-glycerophosphateα-glycerophosphateribonucleotidesglucose6-phosphateFig.
3ContentandpercentageofmajorPtypes(a,b)andmonoestersPcompoundforms(c,d)inalgaecollectedfromsixsites(C1–C6)ofChaoLake169Limnology(2019)20:163–171assumedthatneo-IHPwasnotpresentinalgaeofChaoLake.
DegradationbehaviorsofdiestersPinalgaeofChaoLakeApartfrommonoestersP,otherimportantPocompounds,diestersP,weredetectedinsomealgaesamples.
Thecon-centrationofdiestersPwasgenerallylow(mean0.
63%ofETP),comparedtootherPofractions,andonlydetectedinsamplesC4,C5andC6(9.
2–61.
6mgkg1).
ItiswellestablishedthatsomediestersPsuchasphos-pholipidsandRNAcandegradetomonoestersP[e.
g.
,α-,β-glycerophosphates(phospholipids)andvariousmonophos-phates(e.
g.
,nucleotides)whenanalyzedatthehighpHrequiredforgoodpeakseparationin31PNMRspectra](Turneretal.
2003;Dooletteetal.
2009;Heetal.
2011;Schneideretal.
2016).
ThedegreeofdegradationwillvarydependingonthelengthofNMRexperimentandotherfactors(Cade-MenunandLiu2014;Cade-Menun2015;Fengetal.
2018).
Itwasessentialthatthesedegradationpeakswereidentifiedandquantified,inordertodeterminethecorrectconcentrationsofmonoestersPanddiestersP(Youngetal.
2013;Vincentetal.
2013).
Therefore,thecorrectedtotalmonoestersPandcorrectedtotaldiestersPwerethosecorrectedbymovingthepercentagesofα-andβ-glycerophosphatesandnucleotidesfrommonoestersPtodiestersP.
Whenuncorrected,thetotalmonoestersPweresignificantlyhigherthanthetotaldiestersP,butthereversewastrueforthecorrectedvalues.
EutrophicationandalgalbloomingversusbiogeochemicalcyclingofalgaederivedPinChaoLakeBasedontheresultsinthisstudy,wewereabletothinkfurtheraboutthebiogeochemicalcyclingofPdrivenbyalgalbloominginChaoLake.
TheTPofalgae-derivedbio-massloadingwas10.
94*103kginChaoLake(Lietal.
2015).
WiththePocontentinalgaedeterminedinthisstudy(Table1),weestimatedthePobiomassofalgaetobeapproximately7.
57*103kginChaoLake.
Inpreviousresearch(Fengetal.
2016b)withTaiLakesamples,weesti-matedthatapproximately32.
7–41.
3%ofextractablePofromalgaehasthepotentialforphosphatasehydrolysistosolubleorthophosphatewhichcanbereleasedintothewaterbody.
Fig.
4Solution31PNMRspec-traofmonoestersPstandardcompounds(a–e)andC1algalsampleinChaoLake(f)170Limnology(2019)20:163–171Thus,inthecaseofChaoLake,thisbioavailablePwouldbe2475–3126kginalgaeandwouldbereleasedintothewaterandpromoterepeatedalgalbloomsinChaoLakeifnotappropriatelyremovednaturallyorartificially.
Thiscon-clusionindicatedthatdecompositionofalgaldebriswouldbeakeyfactorinregenerationofbioavailablePforlifeineutrophiclakes,evenwhenexternalPisexcluded.
Itisthere-forenecessarytoremovealgaedebrisfromeutrophiclakestocontrolthereleaseofPfrominternalPcyclingandthephenomenonofeutrophicationoflakes.
ConclusionThisresearchusedsolution31PNMRtoprovideinsightsintothePspeciesanddistributionofPinalgaeoftheheavilypollutedChaoLake.
DataderivedfromthisstudyshowedthattheeutrophiclakealgaehaveaccumulatedremarkableamountsofPiandPo.
TheproportionofPoinalgaerangedfrom55.
3to80.
8%withameanof69.
2%inChaoLake.
EightcompoundsPweredetectedintheNaOH–EDTAextractsofalgalsamplesby31PNMR.
ThesumoforthophosphateandmonoestersPinETPwasgreaterthan93%ofalgalP.
OurobservationsimpliedthatthereleaseofPinducedbythedecompositionofalgaedebriscouldbeapotentialsourceofbioavailablePinaquaticsystemsofChaoLakeevenwithoutanymoreexternalPinput.
Thus,recy-clingofthepotentialbioavailablePinalgaemightbethemechanismofrepeatedalgaebloomingineutrophicChaoLake.
RemediationofthelakerequiresastrategytoremovethealgalbiomassPeffectively.
AcknowledgementsThisresearchwasjointlysupportedbytheNationalNaturalScienceFoundationofChina(41703115,41521003,41630645,41807372)andPostdoctoralScienceFoundationofChina(2017M610967).
OpenAccessThisarticleisdistributedunderthetermsoftheCrea-tiveCommonsAttribution4.
0InternationalLicense(http://creativecommons.
org/licenses/by/4.
0/),whichpermitsunrestricteduse,distribu-tion,andreproductioninanymedium,providedyougiveappropriatecredittotheoriginalauthor(s)andthesource,providealinktotheCreativeCommonslicense,andindicateifchangesweremade.
ReferencesAbdiD,Cade-MenunBJ,ZiadiN,ParentLE(2014)Long-termimpactoftillagepracticesandPfertilizationonsoilPformsasdeter-minedby31P-NMRspectroscopy.
JEnvironQual43:1432–1441BaiX,SunJ,ZhouY,GuL,ZhaoH,WangJ(2017)Variationsofdifferentdissolvedandparticulatephosphorusclassesduringanalgaebloominaeutrophiclakeby31PNMRspectroscopy.
Che-mosphere169:577–585BaldwinDS(2013)Organicphosphorusintheaquaticenvironment.
EnvironChem10:439–454BedrockCN,CheshireMV,ChudekJA,FraserAR,GoodmanBA,ShandCA(1995)EffectofpHonprecipitationofhumicacidfrompeatandmineralsoilsonthedistributionofphosphorusformsinhumicandfulvicacidfractions.
CommunSoilSciPlantAnal26:1411–1425BellDW,PellechiaP,ChambersLR,LongoAF,McCabeKM,IngallED,Benitez-NelsonCR(2017)Isolationandmolecularcharac-terizationofdissolvedorganicphosphorususingelectrodialy-sis-reverseosmosisandsolution31P-NMR.
LimnolOceanogr-Methods.
https://doi.
org/10.
1002/lom3.
10171BusatoJG,CanellasLP,RumjanekVM,VellosoACX(2005)Phos-phorusinaninceptisolunderlong-termsugarcane:II.
HumicacidanalysisbyNMRP-31.
RevBrasCiencSolo29:945–953Cade-MenunBJ(2015)Improvedpeakidentificationin31P-NMRspectraofenvironmentalsampleswithastandardizedmethodandpeaklibrary.
Geoderma257:102–114Cade-MenunBJ,LiuCW(2014)Solutionphosphorus-31nuclearmagneticresonancespectroscopyofsoilsfrom2005to2013:areviewofsamplepreparationandexperimentalparameters.
SoilSciSocAmJ78:19–37Cade-MenunBJ,PrestonCM(1996)Acomparisonofsoilextractionproceduresfor31PNMRspectroscopy.
SoilSci161:770–785ChenY,LiuQ(2014)Onthehorizontaldistributionofalgal-bloominChaohuLakeanditsformationprocess.
ActaMechSinca30:656–666DooletteA,SmernikR,DoughertyW(2009)Spikingimprovedsolu-tionphosphorus-31nuclearmagneticresonanceidentificationofsoilphosphoruscompounds.
SoilSciSocAmJ73:919–927FengW,ZhuY,WuF,MengW,GiesyJP,HeZ,SongL,FanM(2016a)Characterizationofphosphorusformsinlakemacro-phytesandalgaebysolution31Pnuclearmagneticresonancespectroscopy.
EnvironSciPollutRes23:7288–7297FengW,ZhuY,WuF,HeZ,ZhangC,GiesyJP(2016b)Formsandlabilityofphosphorusinalgaeandaquaticmacrophytescharacterizedbysolution31PNMRcoupledwithenzymatichydrolysis.
SciRep6:37164FengW,WuF,HeZ,SongF,ZhuY,GiesyJP,WangY,QinN,ZhangC,ChenH,SunF(2018)Simulatedbioavailabilityofphosphorusfromaquaticmacrophytesandphytoplanktonbyaqueoussuspensionandincubationwithalkalinephosphatase.
SciTotalEnviron616:1431–1439GilesCD,Cade-MenunBJ(2014)Appliedmanureandnutrientchemistryforsustainableagricultureandenvironment.
In:ZhangH(ed)Phytateinanimalmanureandsoils:abundance,cyclingandbioavailability.
Springer,TheNetherland,pp163–190GilesCD,LeeLG,Cade-MenunBJ,HillJE,IslesPD,SchrothAW,DruschelGK(2015)CharacterizationoforganicphosphorusformandbioavailabilityinlakesedimentsusingPnuclearmagneticresonanceandenzymatichydrolysis.
JEnvironQual44:1–13HeZ,HoneycuttCW(2005)Amodifiedmolybdatebluemethodfororthophosphatedeterminationsuitableforinvestigatingenzymatichydrolysisoforganicphosphates.
CommunSoilSciPlantAnal36:1373–1383HeZ,Cade-MenunBJ,ToorGS,SimJT(2007)Comparisonofphos-phorusformsinwetanddriedanimalmanuresbysolutionphos-phorus-31nuclearmagneticresonancespectroscopyandenzy-matichydrolysis.
JEnvironQual36:1086–1095HeZ,HoneycuttCW,Cade-MenunBJ,SenwoZN,TazisongIA(2008)Phosphorusinpoultrylitterandsoil:enzymaticandnuclearmag-neticresonancecharacterization.
SoilSciSocAmJ72:1425–1433HeZ,HoneycuttCW,GriffinTS,Cade-MenunBJ,PellechiaPJ,DouZ(2009)Phosphorusformsinconventionalandorganicdairy171Limnology(2019)20:163–171manureidentifiedbysolutionandsolidstateP-31NMRspectros-copy.
JEnvironQual38:1909–1918HeZ,OlkDC,Cade-MenunBJ(2011)Formsandlabilityofphospho-rusinhumicacidfractionsofHordsiltloamsoil.
SoilSciSocAmJ75:1712–1722JaroschKA,DooletteAL,SmernikRJ,TamburiniF,FrossardE,Büne-mannEK(2015)CharacterisationofsoilorganicphosphorusinNaOH–EDTAextracts:acomparisonof31PNMRspectroscopyandenzymeadditionassays.
SoilBiolBiochem91:298–309KagalouaI,PapastergiadoubE,LeonardasaI(2008)Long-termchangesintheeutrophicationprocessinashallowMediterraneanlakeecosystemofWGreece:responseafterthereductionofexter-nalload.
JEnvironManag87:497–506KarlDM(2014)Microbiallymediatedtransformationsofphosphorusinthesea:newviewsofanoldcycle.
AnnRevMarSci6:279–337LehmanPW,KurobeT,LesmeisterS,BaxaD,TungA,TehSJ(2017)Impactsofthe2014severedroughtontheMicrocystisbloominSanFranciscoestuary.
HarmfulAlgae63:94–108LiM,WuY,YuZ,ShengG,YuH(2009)Enhancednitrogenandphos-phorusremovalfromeutrophiclakewaterbyIpomoeaaquaticawithlow-energyionimplantation.
WaterRes43:1247–1256LiJ,CuiK,LuW,ChengY,JiangY(2015)Communitydynamicsofspring-summerplanktoninLakeChaohu.
ActaHydrobiolSinica39:185–194(inChinese)LiuS,ZhuY,MengW,HeZ,FengW,ZhangC,GiesyJP(2016)Char-acteristicsanddegradationofcarbonandphosphorusfromaquaticmacrophytesinlakes:insightsfromsolid-state13CNMRandsolu-tion31PNMRspectroscopy.
SciTotalEnviron543:746–756LiuS,ZhuY,WuF,MengW,WangH,HeZ,GuoW,SongF,GiesyJP(2017)Usingsolid13CNMRcoupledwithsolution31PNMRspectroscopytoinvestigatemolecularspeciesandlabilityoforganiccarbonandphosphorusfromaquaticplantsinTaiLake.
ChinaEnvironSciPollutRes24:1880–1889MahieuN,OlkDC,RandallEW(2000)Analysisofphosphorusintwohumicacidfractionsofintensivelycroppedlowlandricesoilsby31PNMR.
EurJSoilSci51:391–402McDowellRW,HillSJ(2015)Speciationanddistributionoforganicphosphorusinriversediments:anationalsurvey.
JSoilSediment15:2369–2379Pernet-CoundrierB,QiWX,LiuHJ,NüllerB,BergM(2012)Sourcesandpathwayofnutrientsinthesemi-aridregionofBeijing–Tian-jin.
ChinaEnvironSciTechnol46:5294–5301QuX,XieL,LinY,BaiY,ZhuY,XieF,GeisyJP,WuF(2013)Quan-titativeandqualitativecharacteristicsofdissolvedorganicmatterfromeightdominantaquaticmacrophytesinLakeDianchi.
ChinaEnvironSciPollutRes20:7413–7423ReitzelK,AhlgrenJ,GogollA(2006)Effectsofaluminumtreatmentonphosphorus,carbon,andnitrogendistributioninlakesediment:a31PNMRstudy.
WaterRes40:647–654RubanV,López-SánchezJF,PardoP(2001)Harmonizedprotocolandcertifiedreferencematerialforthedeterminationofextractablecontentsofphosphorusinfreshwatersediments—asynthesisofrecentworks.
FresenJAnalChem370:224–228SchneiderK,ReslP,SpribilleT(2016)Escapefromthecrypticspeciestrap:lichenevolutiononbothsidesofacyanobacterialacquisitionevent.
MolEcol25:3453–3468SmernikRJ,DoughertyWJ(2007)Identificationofphytateinphos-phorus-31nuclearmagneticresonancespectra:theneedforspik-ing.
SoilSciSocAmJ71:1045–1050SrensenDR,NielsenUG,SkouEM(2014)Solidstate31PMASNMRspectroscopyandconductivitymeasurementsonNbOPO4andH3PO4compositematerials.
JSolidStateChem219:80–86SuzumuraM(2005)Phospholipidsinmarineenvironments:areview.
Talanta66:422–434TangJ,ShiT,WuX,CaoH,LiX,HuaR,TangF,YueY(2015)TheoccurrenceanddistributionofantibioticsinLakeChaohu,China:seasonalvariation,potentialsourceandriskassessment.
Chem-osphere122:154–161TurnerBL,RichardsonAE(2004)Identificationof-inositolphosphatesinsoilbysolutionphosphorus-31nuclearmagneticresonancespectroscopy.
SoilSciSocAmJ68:802–808TurnerBL,MahieuN,CondronLM(2003)Phosphorus-31nuclearmagneticresonancespectralassignmentsofphosphoruscom-poundsinsoilNaOH–EDTAextracts.
SoilSciSocAmJ67:497–510TurnerBL,Cade-MenunBJ,CondronLM,NewmanS(2005)Extrac-tionofsoilorganicphosphorus.
Talanta66:294–306TurnerBL,CheesmanAW,GodageHY,RileyAM,PotterBVL(2012)Determinationofneo-andd-chiro-inositolhexakisphosphateinsoilsbysolution31PNMRspectroscopy.
EnvironSciTechnol46:4994–5002VincentAG,VestergrenJ,GrbnerG,PerssonP,SchleucherJ,BieslerR(2013)Soilorganicphosphorustransformationsinaborealfor-estchronosequence.
PlantSoil367:149–162WangJ,PantHK(2010)Enzymatichydrolysisoforganicphosphorusinriverbedsediments.
EcoEng36:963–968WangX,XiB,HuoS,DengL,PanH,XiaX,ZhangJ,RenY,LiuH(2013)PolybrominateddiphenylethersoccurrenceinmajorinflowingriversofLakeChahu(China):characteristics,potentialsourcesandinputstolake.
Chemosphere93:1624–1631XuM,CaoH,XieP,DengD,FengW,XuJ(2005)Thetemporalandspatialdistribution,compositionandabundanceofprotozoainChaohuLake,China:relationshipwitheutrophication.
EuropJProtistol41:183–192YangL,HanX,SunP,YanW,LiY(2011)CanonicalcorrespondenceanalysisofalgaecommunityanditsenvironmentalfactorsinLakeChaohu.
ChinaJAgro-EnvironSci30:952–958YoungEO,RossDS,Cade-MenunBJ,LiuCW(2013)Phosphorusspeciationinripariansoils:aphosphorus-31nuclearmagneticresonancespectroscopyandenzymehydrolysisstudy.
SoilSciSocAmJ77:1636–1647ZanF,HuoS,XiB,LiQ,LiaoH,ZhangJ(2010)Phosphorusdistribu-tioninthesedimentsofashalloweutrophiclake.
LakeChaohu,China.
EnvironEarthSci62:1643–1653ZhangWQ,XingBS(2013)DetectionofphosphorusspeciesinthesedimentsofChaohuLakeby31Pnuclearmagneticresonancespectroscopy(31P-NMR).
ActaSciCircum33:1967–1973(inChinese)ZhangWQ,ZhuXL,JinX,MengX,TangWZ,ShanBQ(2017)Evi-dencefororganicphosphorusactivationandtransformationatthesediment-waterinterfaceduringplantdebrisdecomposition.
SciTotalEnviron583:458–465ZhongW,ZhangZ,LuoY(2012)Biogasproductivitybyco-digestingTaihubluealgaewithcornstrawasanexternalcarbonsource.
BioresourceTechnol114:281–286ZhuG,QinB,ZhangL,LuoL(2006)Geochemicalformsofphos-phorusinsedimentsofthreelarge,shallowlakesofChina.
Pedo-sphere16:726–734ZhuY,WuF,HeZ,GuoJ,QuX,XieF,GiesyJP,LiaoH,GuoF(2013)Characterizationoforganicphosphorusinlakesedimentsbysequentialfractionationandenzymatichydrolysis.
EnvironSciTechnol47:7679–7687ZhuY,WuF,HeZ(2015)Bioavailabilityandpreservationoforganicphosphorusinfreshwatersedimentsanditsroleinlakeeutrophi-cation.
In:HeZ,WuF(eds)Labileorganicmatter—chemicalcompositions,function,andsignificanceinsoilandtheenviron-ment.
SSSA,Madison,pp275–294ZhuY,WuF,FengW,LiuS,GiesyJP(2016)Interactionofalkalinephosphatasewithmineralsandsediments:activities,kineticsandhydrolysisoforganicphosphorus.
ColloidssurfA:PhysicochemEngAspects495:46–53
org/10.
1007/s10201-018-0562-2RESEARCHPAPERCharacterizationofphosphorusinalgaefromaeutrophiclakebysolution31PnuclearmagneticresonancespectroscopyWeiyingFeng1·CuicuiLi1,2,3·ChenZhang1·ShashaLiu1·FanhaoSong1·WenjingGuo1·ZhongqiHe4·TingtingLi1·HaiyanChen1Received:4July2018/Accepted:12September2018/Publishedonline:4October2018TheAuthor(s)2018AbstractTheidentificationandquantificationofphosphorus(P)compoundsderivedfromalgalbiomassarecrucialforabetterunderstandingofalgalPdynamicsinlakeecosystems.
QuantityandspeciesofPinalgaecollectedfromChaoLake(atypi-calultra-eutrophiclake)inChinawereanalyzedbychemicalanalysisand31PNMR.
TotalP(TP)inalgaebiomassrangedfrom2671to5385mgkg1ofdrymatter.
ProportionoforganicP(Po)accountedfor78.
3±2.
6%inalgaebiomasscollectedfromthewesternpartofChaoLake,whichwashigherthanthat(64.
7±1.
4%)intheeasternpartofthelake.
EightPspeciesincludinginorganicPspecies(orthophosphateandpyrophosphate)andPospecies(fivemonoestersPanddiestersP)wereidentifiedinNaOH–EDTAextractsofalgalsamples.
MonoestersPaccountedfor48.
4%inextractedTP,whichwasthemaincomponentofPo.
β-glycerophosphateswerethelargestcomponentofmonoestersP,whichaccountedfor22.
6%inextractedTP.
ThisstudyimprovedknowledgeonthemechanismofthecyclingofendogenousPintheaquaticsystemandwouldbehelpfulindevelopingastrategyforcontrolofrepeatedalgaebloomsineutrophicChaoLake.
Keywords31PNMR·Algae·Organicphosphorus·Lake·EutrophicationIntroductionAlgaebloomsareseriousenvironmentalproblemsaroundtheworld,especiallyindevelopingcountries(Kagalouaetal.
2008;Pernet-Coundrieretal.
2012).
Algaebloomsoccurinanaquaticenvironmentiftoomuchphosphorus(P)entersthesystem(Gilesetal.
2015).
Foraeutrophiclake,releaseofPfromdeadalgaeisanimportantnutrientsourcethatwillsupportcontinuousalgalbloomsinlakes(Lietal.
2009;Fengetal.
2016a).
Algaearenotusuallycollectedbyhumansbecausetheyarenotvaluable,sothatthedebrisisallowedtodecomposeinsitu.
Thedecomposi-tionofalgalresiduesaffectsthebio-cyclingandreleaseofP,increasingtheriskofresurgenceofalgalblooms(Fengetal.
2016b;Lehmanetal.
2017).
DecayingalgaldebrisreleasesbothinorganicP(Pi)andPo.
ThePoconstituentsneedtobehydrolyzedtobioavailablePobyvariousenzymes(Fengetal.
2016b).
Therefore,theformsandconcentrationsofPinalgaeshallbeevaluatedwhilealgaedebrisisdecomposing.
However,untilnow,studiesofthespecies,concentrations,andeffectsofPoinalgaeofeutrophiclakeshavebeenlim-itedbecauseofthecomplexityandlimitationsofanalyticalmethods(Turneretal.
2005;Belletal.
2017).
Typicalanalyticalapproaches,suchasenzymaticreac-tivity,highperformanceliquidchromatography(HPLC),andmassspectrometryarebasedonoperationaldefini-tionssothattheycannotdiscernPclassesatamolecularlevel(Suzumura2005;Baldwin2013;Karl2014).
Phos-phorus-31nuclearmagneticresonancespectroscopy(31PHandlingEditor:RichardSheibley.
WeiyingFengandCuicuiLiareco-leadauthors.
*ChenZhangzhangchen0353@yeah.
net*HaiyanChenchenhy@craes.
org.
cn1StateKeyLaboratoryofEnvironmentalCriteriaandRiskAssessment,ChineseResearchAcademyofEnvironmentalSciences,Beijing100012,China2GuangzhouInstituteofGeochemistry,ChineseAcademyofSciences,Guangzhou510640,China3UniversityofChineseAcademyofSciences,Beijing100049,China4USDAARS,SouthernRegionalResearchCenter,NewOrleans,LA70124,USA164Limnology(2019)20:163–171NMR)isanon-destructive,non-invasivetechniqueforiden-tifyingchemicalformsinvariousenvironmentalsamples.
Solid-state31PNMRisfocusedoninorganicPcompounds,solution31PNMRismainlyusedtodetermineorganicPcompounds(Turneretal.
2005;Heetal.
2011;Abdietal.
2014;Srensenetal.
2014).
SeveralPcompoundshavebeendetectedbysolution31PNMR,includingphosphonates,orthophosphate,monoestersP,diestersP,pyrophosphatesandpolyphosphates(Heetal.
2007;Turneretal.
2012;Zhuetal.
2013).
Generally,monoestersPrepresentawiderangeofimportantPocompounds,suchasinositolphos-phateandsugarphosphates(Heetal.
2007;Dooletteetal.
2009;Jaroschetal.
2015).
Therefore,itisanidealtechniqueforanalyzingPospeciesinalgaeofeutrophiclakes,asitwouldnotonlyprovideimportantinformationpertainingtoPbiogeochemicalcyclinginlakeecosystems,butalsoyieldabundantinsightintoidentitiesofspecificPcompounds.
ChaoLake(31°25′28″–31°43′28″N,117°16′54″–117°51′46″E)isoneofthefivelargestfreshwa-terlakesinChina.
ItissituatedonthefloodplainsbetweentheYangtzeRiverandHuaiRiverinthecentralAnhuiProv-inceofeasternChina(Zanetal.
2010;Tangetal.
2015).
ChaoLakeisatypicalshallowlakewithameandepthof3mandasurfaceareaof780km2anddrainageareaof13,350km2(Wangetal.
2013).
Duetotherapidincreaseinanthropogenicactivitiesinthelake'swatershedoverrecentdecades,thelakehassufferedfromseriouspollution,eutrophicationandalgaeblooms(Xuetal.
2005).
Asamat-teroffact,sincethemid1980s,algaebloomshaveoccurredeachyearinChaoLake(ChenandLiu2014).
Inordertoassesstheenvironmentalrisksofalgaeontheeutrophiclake,thisstudyanalyzedthePspeciesinalgaebysolution31PNMR,andbasedontheknowledgeobtained,predictedthePbioavailabilityofthealgalbiomassinaquaticenvironments.
MaterialsandmethodsStudysitesandsamplecollectionSamplesofalgaewerecollectedfromsixsitesinChaoLakeinSeptember2015(Fig.
1).
Thesesamplingsiteswerelocatedindifferenteutrophicareas.
Generally,ChaoLakeisdividedintotwoparts:thewesternpart(samplesC1andC2)andtheeasternpart(samplesC3,C4,C5andC6)alongthelineofZhongmiao–Mushan–Qitouzui,asshowninFig.
1.
Thequalityofwaterwasworstinthewesternpartofthelakeandgraduallybecamebetterfromwesttoeast(Zhuetal.
Fig.
1MajorinletsandoutletsofChaoLakewithalgalsamplingsites165Limnology(2019)20:163–1712006;Tangetal.
2015).
Thelake'sannualmeanconcentra-tionsoftotalnitrogen(TN)andTPapproached2.
85and0.
26mgl1,respectively,andtheannualmeanchlorophyll-areachedupto25.
6μgl1(Lietal.
2015).
Algaewerecol-lectedbyuseofaplanktoncollector(HB403-BWS).
Sam-pleswereplacedinsealedbagsandputiniceboxesimme-diately.
Thesealgalsampleswerefreeze-dried,ground,thenpassedthrougha2-mmsievebeforebeingstoredat20°C(Fengetal.
2016a).
ThedominantspeciesinChaoLakewasMicrocysticaeruginosawithanappearancefrequencyof90.
9%(Yangetal.
2011).
ExtractionofPandchemicalanalysisSamplesofalgaewereextractedbyuseofopti-mizedNaOH–EDTAextractingagent(mixturesof0.
5moll1NaOHand25mmoll1EDTA)witharatioof150:1(mlg1),andthemixtureswereshakenfor18hatroomtemperature(Cade-MenunandPreston1996;Fengetal.
2016b).
Theextractingsolutionswerecentrifuged(8000*g)for30min,andfilteredthrough0.
45-μmglass-fiberfilters(WhatmanGF/C).
Extractabletotalphosphorus(ETP)afterdigestionandfreemolybdatereactivephos-phorus(MRP)weremeasuredusingthemolybdenumbluemethod(HeandHoneycutt2005).
Extractableorganicphos-phorus(EOP)wascalculatedbythedifferencebetweenETPandMRP.
Theremainingextractswerefreeze-driedforsolu-tion31PNRMspectroscopyanalysis.
Percentagesofcarbon(C)andnitrogen(N)inalgaeweredeterminedbyuseofanelementalanalyzer(ElementarvariomacroEL,Germany).
Totalphosphorus(TP)andinorganicphosphorus(Pi)weredeterminedbytheSMTmethoddescribedbyRubanetal.
(2001).
Organicphosphorus(Po)inalgaesampleswascalculatedbythedifferencebetweenTPandPi.
TherewerethreereplicationsforextractionofPandchemicalanalysis.
Datawerecheckedfordeviationsfromnormalityandhomogeneityofvariancebeforeperformingstatisticalanalyses.
31PNMRanalysisA100-mgsampleoffreeze-driedalgalextractswasground,andthenredissolvedin1ml1moll1NaOH+0.
1moll1EDTAand0.
2mlD2O.
Afterultrasonicationfor30minandequilibrationfor5min,2%(v/v)ofbicarbonatebuffereddithionite(0.
11moll1NaHCO3+0.
11moll1Na2S2O4)wasaddedtotheextracttoreduceinterferencefrompara-magneticions,suchasFeandMn(Heetal.
2009;GilesandCade-Menun2014).
ThepHofthesupernatantsolutionwasadjustedusing10moll1NaOHtoensureapH>12.
Thesupernatantsolutionwascentrifuged(8000*g)for30minandtransferredtoa5-mmNMRtube.
Solution31PNMRspectrawereacquiredat24°ConaBrukerAVANCE400MHzspectrometerata31Pfrequencyof161.
98MHz,usinga90°pulse,a5srelaxationdelayanda0.
21sacqui-sitiontime,similartotheparametersusedinFengetal.
(2016b).
Thescantimeforeachsamplewasmorethan15h.
PeakareaswerecalculatedbyintegrationandcompletedusingMestrelabMNovav.
10.
SpikingexperimentsThepeakofspecificmonoestersPforms(i.
e.
,glu-cose6-phosphate,riboncleotides,α-glycerophosphate,β-glycerophosphate,myo-inositolhexaphosphate)needstobeconfirmedwithspikingexperiments(Fig.
4).
MethodsforidentifyingspecificPformsinNMRspectraofsoilandotherenvironmentalsamplesarewell-establishedandhavebeenusedformanyyears(SmernikandDougherty2007;Heetal.
2011;McDowellandHill2015),combinedwithPcom-poundlibrariesdevelopedbyTurneretal.
(2003);Dooletteetal.
(2009)andCade-Menun(2015).
StandardsamplesofmonoestersPwerepurchasedfromSigma-Aldrich.
Spikedsampleswereanalyzedby31PNMRasdescribedabove.
MonoestersPcompoundswereidentifiedbytheirchemicalshifts,withtheorthophosphatepeakinallspectrastand-ardizedto6.
0ppm.
SpectralprocessingwasdoneusingMestReNovesoftwareversion9.
0.
1(MestReNoveResearchSL).
ResultsandanalysisNutrients(C,NandP)indebrisofalgaeCompositionofC,NandtheirratiosindebrisofalgaeareshowninTable1.
PercentagesofCrangedfrom31.
5to50.
3%withameanvalueof41.
9%inthealgaesamplesfromChaoLake.
ContentofNwas5.
2–7.
8%withameanvalueof6.
4%.
BothcontentsofCandNofalgaewasgreaterthanthoseofaquaticmacrophytes,whichhasbeenwidelyreportedpreviously(Zhongetal.
2012;Quetal.
2013,Fengetal.
2016a).
TheratioofC:Nwasagoodpredictorofdeg-radationinalgaeandaquaticmacrophyteswithalowerratioofC:Nformaterialmorereadilydegradation(Reitzeletal.
2006).
Inthisstudy,theratiosofC:Ninalgaefromthewest-ernlake(7.
1±0.
3)werehigherthanthoseoftheeasternlake(6.
4±0.
3),suggestinghigherlabilityofthealgaldebrisoftheeasternlake.
TheratioofC:Nofaquaticmacrophytes(average12.
9)(Fengetal.
2016a)wasgreaterthanthatofalgaeinChaoLake(average6.
7)(Table1).
Therefore,thealgaedecomposedmoreeasilythanaquaticmacrophytesinthesamelake.
Thiswasalsoconsistentwiththeresultsofpreviousstudies(Liuetal.
2016).
ContentsofTPindebrisofalgaerangedfrom2671to5385mgkg1drymass(dm)withameanvalueof166Limnology(2019)20:163–1713936mgkg1.
ThegreatestconcentrationofTPindebrisofalgaewasobservedinsampleC6.
Thismaybebecauseoftwomajorinflowingrivers(e.
g.
,theZhegaoriverandShuangqiaoriver)withheavynutritioninputs(Tangetal.
2015).
ContentsofTPinthesurfacesedimentsfromChaoLakeonlyrangedfrom420to1090mgkg1withameanvalueof687mgkg1(ZhangandXing2013).
ContentofTPindebrisofalgaefromChaoLakewasapproximately5timeshigherthanTPinsurfacesediments.
Therefore,deadalgae-derivedPmightbeanimportantsourceofbioavail-ablePforrepeatedalgalbloomingineutrophiclakessuchasChaoLake.
ConcentrationofPointhesealgalsamplesrangedfrom2107to4136mgkg1withameanvalueof2783mgkg1.
ThegreatestconcentrationofPowasalsoobservedintheheavilypollutedregion(i.
e.
,sampleC6).
TheproportionofPoinalgaeofthewesternlake(78.
3%±2.
6%)washigherthanthatintheeasternlake(64.
7%±1.
4%).
ThemeanratioofPo/TPofthesixalgalsampleswas69.
2%inChaoLake(Table1).
ThisvaluewasgreaterthanthatofalgaefromTaiLake(meanratioofPo/TP57.
8%)(Fengetal.
2016b).
ThepreviousstudiesreportedthatPocouldbecon-vertedtobioavailableP(e.
g.
,HPO42)foralgaebloomingthroughaseriesofredox-drivensolubilizationreactionsandphosphatase-mediatedhydrolyticprocesses(WangandPant2010;Zhuetal.
2015).
Thus,webelievedthatPoindebrisofalgaefromChaoLakepossessedlargerbioavailabilitypotentialthanalgaefromTaiLakepertheirdifferenceinPo/TPratios.
NaOH–EDTAextractablePfromalgaeInthisstudy,contentsofNaOH–EDTAextractableTPrangedfrom1292to2774mgkg1,withanextractionefficiencyof40.
5–78.
3%,andcontentsofextractablePofromalgaerangedfrom351to1141mgkg1,withanextrac-tionefficiencyof13.
8–54.
2%(Table1).
TherecoveriesofPoindebrisofalgaefromChaoLakewerelowerthanthosewithpurealgaesuchasMicrocystis,Chlorellavulgaris,andSprilinaplatensis(Fengetal.
2016a).
However,theextrac-tionefficienciesofPoofalgaefromChaoLakeweresimilartothoseofparticulatePfromTaiLake(23–56%)(Baietal.
2017)andofsedimentsfromHaiheRiver(30–73%)(Zhangetal.
2017).
TheextractionefficiencyofTPfromalgaefromChaoLakewassimilartothatofsoilsandsediments(49–83%)(Xuetal.
2005;Tangetal.
2015).
Multiple-stepextractions(e.
g.
,additionalorsequentialHClextraction)(Heetal.
2008;Cade-Menun2015;Zhuetal.
2016;Liuetal.
2017)seemneededtoincreasethePrecoveryfromthesealgalsamples.
Solution31PNMRspectraofNaOH–EDTAextractsofalgaeEightmainPspeciesincludinginorganicPspecies(orthophosphateandpyrophosphate)andPospecies(fivemonoestersPanddiestersP)wereidentifiedintheNaOH–EDTAextractsofthesixalgalsamplesbysolution31PNMR(Fig.
2;Table2).
Thepeakoforthophosphatewasat6.
00ppminthe31PNMRspectra,monoestersPwasat3.
33–5.
49ppm,diestersPwasat0.
69to0.
31ppm,andpyrophosphatewasat4.
21to4.
12ppm(Fig.
2).
ThesumoforthophosphateandmonoestersPinETPaccountedformorethan93%ofETP(Table1;Fig.
3b).
Withthepeakoforthophosphateasthelargestsignalinthese31PNMRspectra.
Thecontentoforthophosphatewasbetween615.
1and1331.
7mgkg1,andaccountedTable1ContentsofC,N,andPinalgaeandtheirNaOH–EDTAextractionefficiencyinChaoLakeETPextractabletotalphosphorus,EOPextractableorganicphosphorusaMean±standarddeviation(n=3)bValuesinbracketsshowthepercentageofETP,EOPinNaOH–EDTAextractstoTPandPointheunextractedalgaepowders,respectivelySamplesCoordinatesOriginalalgaepowdersNaOH–EDTAextractalgaesamplesC(%)N(%)C:NTP(mgkg1)Po(mgkg1)Po/TP(%)ETP(mgkg1)EOP(mgkg1)C131°38′3.
52″E,117°21′16.
74″N44.
8±2.
5a6.
7±0.
56.
7±0.
34173±5163372±12680.
8±1.
61864±126(44.
7)b785±124(23.
3)bC231°34′3.
50″E,117°24′51.
80″N39.
5±0.
65.
3±0.
87.
5±0.
24059±1253072±43275.
7±3.
51757±214(43.
3)464±52(15.
1)C331°30′9.
64″E,117°28′56.
52″N38.
3±1.
85.
4±0.
27.
1±0.
43787±2562536±5967.
0±2.
51572±56(41.
5)351±98(13.
8)C431°27′56.
84″E,117°34′50.
00″N31.
5±3.
55.
2±0.
16.
1±0.
12671±1981476±12355.
3±1.
41292±89(48.
4)567±15(38.
4)C531°33′27.
70″E,117°36′46.
18″N50.
3±2.
97.
8±0.
86.
5±0.
43543±1122107±7859.
5±0.
92774±21(78.
3)1141±215(54.
2)C631°36′10.
25″E,117°47′38.
55″N46.
9±4.
67.
8±0.
76.
0±0.
25385±2904136±1976.
8±0.
82183±51(40.
5)1066±164(25.
8)167Limnology(2019)20:163–171for41.
5–54.
0%ofETP(Table1;Fig.
3).
PolyphosphateswerenotdetectedinanyalgaeofChaoLake.
MonoestersPcomprisedthelargestPofractionwithNaOH–EDTAextracts,andaccountedfor48.
4%(average)inETP(Table2;Fig.
3b).
Throughthespikingexperi-ments(Fig.
4),thepeakat4.
88±0.
02ppmwasassignedtoα-glycerophosphatesandthepeakat4.
50±0.
02ppmwasassignedtoβ-glycerophosphates,basedonTurnerandRichardson(2004)andDooletteetal.
(2009);thepercentagesofα-andβ-glycerophosphatesinETPwere3.
4%(65.
5mgkg1)and22.
6%(432.
7mgkg1),respec-tively.
β-glycerophosphateswerethelargestcomponentofmonoestersP.
Thepeakat4.
32±0.
01ppmwasassignedtoribonucleotides(Heetal.
2011)andthepercentageofribonucleotidesinETPwas10.
6%(218.
2mgkg1).
Thepeakat5.
12±0.
01ppmwasassignedtoglucose6-phos-phate(Cade-Menun2015),thepercentageofwhichinETPwas1.
9%(38.
7mgkg1).
Inaddition,afewofthepeaksinthemonoestersPregionwereunidentified,becausechemicalshiftswerestronglyinfluencedbysubtlediffer-encesamongsamplesforviscosity,pH,saltsandparamag-neticions(Youngetal.
2013;Abdietal.
2014;Heetal.
2011;Gilesetal.
2015).
UnidentifiedmonoestersParedefinedas'othermonoestersP'inthisstudy.
TheseothermonoestersPaccountedfor9.
9%ofETPinalgaeofChaoLake(Table2).
PyrophosphatewasdetectedinmostsamplesexceptthesampleC1,whichisconsistentwithanumberofotherstud-ies(Bedrocketal.
1995;Mahieuetal.
2000;Heetal.
2011).
Inotherliterature,polyphosphatesandpyrophosphatewere6420-2-4C2C1OrthophosphatePyrophosphateMonoestersPC3^C5C4^C6DiestersPChemicalshift(ppm)Fig.
2Solution31PNMRspectraofNaOH–EDTAextractsofalgaeinChaoLakeTable2ConcentrationsofPcompoundsinNaOH–EDTAextractsofthealgaedeterminedbysolution31PNMRPiinorganicP,PoorganicP,TotalPothesumofmonoestersPanddiestersP,n.
dnotdetectedaValuesinparenthesesarepercentagesofindividualPcompoundsinNaOH–EDTAextractsTPAlgaePi(mgkg1)Po(mgkg1)OrthophosphatePyrophosphateMonoestersPDiestersPTotalPoGlucose6-phos-phateRibonucleotidesα-glycerophosphateβ-glycerophosphateOthermonoestersPTotalmonoestersPC1818.
2(43.
9)an.
d32.
7(1.
8)204.
6(11.
0)60.
6(3.
2)526.
9(28.
3)220.
9(11.
9)1045.
7(56.
1)n.
d1045.
7(56.
1)C2938.
1(53.
4)115.
4(6.
6)42.
2(2.
4)144.
5(8.
2)63.
8(3.
6)322.
7(18.
4)130.
4(7.
4)703.
5(40.
0)n.
d703.
5(40.
0)C3849.
1(54.
0)15.
8(1.
0)n.
d137.
6(8.
7)49.
2(3.
1)325.
2(20.
5)201.
5(12.
7)714.
1(45.
0)n.
d714.
1(45.
0)C4615.
1(47.
6)65.
8(5.
1)15.
4(1.
2)131.
2(8.
8)44.
9(3.
5)297.
7(23.
0)131.
0(10.
1)604.
4(46.
6)9.
2(0.
7)611.
4(47.
3)C51331.
7(48.
0)87.
9(3.
2)49.
3(1.
8)487.
4(17.
6)103.
9(3.
7)636.
6(22.
9)67.
9(2.
4)1346.
1(48.
5)9.
3(0.
3)1354.
4(48.
8)C6905.
6(41.
5)32.
6(1.
5)92.
4(4.
2)203.
8(9.
3)70.
6(3.
2)487.
2(22.
3)329.
6(15.
1)1184.
0(54.
2)61.
6(2.
8)1245.
2(57.
0)168Limnology(2019)20:163–171detectedinsomesamples,butnotnecessarily(Busatoetal.
2005;Fengetal.
2016b).
DiscussionIdentificationofinositolhexaphosphate(IHP)inalgaeofChaoLakeTheinositolhexaphosphate(IHP)stereoisomers(scyllo-,myo-,chiro-,neo-IHP)wereimportantmonoestersPcom-ponentsinmanyenvironmentalsamples(Turneretal.
2012;Cade-Menun2015).
Eachofthesecompoundscontainssixphosphates,andtheconformationofthosephosphategroupscausesthemtohavemultiplepeaksinasinglespectrum,inanarrangementspecifictoeachcom-pound.
Theonlyexceptionisscyllo-IHP,whichhasonepeakforthesixphosphates;thepeakat3.
55±0.
02ppmwasassignedtoscyllo-IHP,basedonTurnerandRichard-son(2004)andDooletteetal.
(2009).
NoneofthespectrashowninFig.
2haveapeakat3.
55ppm,sothescyllo-IHPwasnotpresentinalgaeofChaoLake.
Myo-IHPhasfourpeaksina1:2:2:1arrangement(withrespecttopeakareas),withpeaksat5.
79±0.
01,4.
88±0.
01,4.
55±0.
01and4.
42±0.
01ppm(inFig.
4e).
ThreeofthesepeakswerenotpresentinalgaesamplesinChaoLake(Fig.
2),somyo-IHPwasnotpresentinalgaeofChaoLake.
Thefactthatmyo-IHPhadnotbeenobservedinalgaeinapreviousstudyisconsistentwiththeobservationofthisstudy(Fengetal.
2016a).
Forchiro-IHP[ineitherthe4equatorial/2axial(4e/2a)conformationorthe2e/4aconformation],threepeaksmustbeclearlyvisible,ina2:2:2arrange-ment,andthediagnosticpeaksforeachareat6.
2–6.
5ppm(Cade-Menun2015);however,noneofthespectrashowninFig.
2haveanypeaksbetween6.
0and7.
0ppm,sothesecompoundswerenotpresentinalgaeofChaoLake.
Inaddition,neo-IHPrequirestwopeakstobepresent,ina4:2arrangement,sothatthepeakat6.
4±0.
01ppmistwiceaslargeastheoneat4.
3±0.
01ppm.
Giventhattherewerenopeaksbetween6.
0and7.
0ppm,itcanbe050010001500200025003000C1C2C3C4C5C6&RQFHQWRIPDMRU3W\SHVPJ3NJ$OJDOVDPSOHV(a)(b)(c)(d)pyrophosphatediestersPmonoestersPorthophosphate0%20%40%60%80%100%C1C2C3C4C5C63HUFHQWDJHRIPDMRU3W\SHVLQ(73$OJDOVDPSOHVorthophosphatemonoestersPdiestersPpyrophosphate0200400600800100012001400C1C2C3C4C5C6&RQFHQWRIPRQRHVWHUV3$OJDOVDPSOHVOthermonoestersPβ-glycerophosphateα-glycerophosphateribonucleotidesglucose6-phosphate0%20%40%60%80%100%C1C2C3C4C5C63HUFHQWDJHRIPRQRHVWHUV3LQ(73$OJDOVDPSOHVOthermonoestersPβ-glycerophosphateα-glycerophosphateribonucleotidesglucose6-phosphateFig.
3ContentandpercentageofmajorPtypes(a,b)andmonoestersPcompoundforms(c,d)inalgaecollectedfromsixsites(C1–C6)ofChaoLake169Limnology(2019)20:163–171assumedthatneo-IHPwasnotpresentinalgaeofChaoLake.
DegradationbehaviorsofdiestersPinalgaeofChaoLakeApartfrommonoestersP,otherimportantPocompounds,diestersP,weredetectedinsomealgaesamples.
Thecon-centrationofdiestersPwasgenerallylow(mean0.
63%ofETP),comparedtootherPofractions,andonlydetectedinsamplesC4,C5andC6(9.
2–61.
6mgkg1).
ItiswellestablishedthatsomediestersPsuchasphos-pholipidsandRNAcandegradetomonoestersP[e.
g.
,α-,β-glycerophosphates(phospholipids)andvariousmonophos-phates(e.
g.
,nucleotides)whenanalyzedatthehighpHrequiredforgoodpeakseparationin31PNMRspectra](Turneretal.
2003;Dooletteetal.
2009;Heetal.
2011;Schneideretal.
2016).
ThedegreeofdegradationwillvarydependingonthelengthofNMRexperimentandotherfactors(Cade-MenunandLiu2014;Cade-Menun2015;Fengetal.
2018).
Itwasessentialthatthesedegradationpeakswereidentifiedandquantified,inordertodeterminethecorrectconcentrationsofmonoestersPanddiestersP(Youngetal.
2013;Vincentetal.
2013).
Therefore,thecorrectedtotalmonoestersPandcorrectedtotaldiestersPwerethosecorrectedbymovingthepercentagesofα-andβ-glycerophosphatesandnucleotidesfrommonoestersPtodiestersP.
Whenuncorrected,thetotalmonoestersPweresignificantlyhigherthanthetotaldiestersP,butthereversewastrueforthecorrectedvalues.
EutrophicationandalgalbloomingversusbiogeochemicalcyclingofalgaederivedPinChaoLakeBasedontheresultsinthisstudy,wewereabletothinkfurtheraboutthebiogeochemicalcyclingofPdrivenbyalgalbloominginChaoLake.
TheTPofalgae-derivedbio-massloadingwas10.
94*103kginChaoLake(Lietal.
2015).
WiththePocontentinalgaedeterminedinthisstudy(Table1),weestimatedthePobiomassofalgaetobeapproximately7.
57*103kginChaoLake.
Inpreviousresearch(Fengetal.
2016b)withTaiLakesamples,weesti-matedthatapproximately32.
7–41.
3%ofextractablePofromalgaehasthepotentialforphosphatasehydrolysistosolubleorthophosphatewhichcanbereleasedintothewaterbody.
Fig.
4Solution31PNMRspec-traofmonoestersPstandardcompounds(a–e)andC1algalsampleinChaoLake(f)170Limnology(2019)20:163–171Thus,inthecaseofChaoLake,thisbioavailablePwouldbe2475–3126kginalgaeandwouldbereleasedintothewaterandpromoterepeatedalgalbloomsinChaoLakeifnotappropriatelyremovednaturallyorartificially.
Thiscon-clusionindicatedthatdecompositionofalgaldebriswouldbeakeyfactorinregenerationofbioavailablePforlifeineutrophiclakes,evenwhenexternalPisexcluded.
Itisthere-forenecessarytoremovealgaedebrisfromeutrophiclakestocontrolthereleaseofPfrominternalPcyclingandthephenomenonofeutrophicationoflakes.
ConclusionThisresearchusedsolution31PNMRtoprovideinsightsintothePspeciesanddistributionofPinalgaeoftheheavilypollutedChaoLake.
DataderivedfromthisstudyshowedthattheeutrophiclakealgaehaveaccumulatedremarkableamountsofPiandPo.
TheproportionofPoinalgaerangedfrom55.
3to80.
8%withameanof69.
2%inChaoLake.
EightcompoundsPweredetectedintheNaOH–EDTAextractsofalgalsamplesby31PNMR.
ThesumoforthophosphateandmonoestersPinETPwasgreaterthan93%ofalgalP.
OurobservationsimpliedthatthereleaseofPinducedbythedecompositionofalgaedebriscouldbeapotentialsourceofbioavailablePinaquaticsystemsofChaoLakeevenwithoutanymoreexternalPinput.
Thus,recy-clingofthepotentialbioavailablePinalgaemightbethemechanismofrepeatedalgaebloomingineutrophicChaoLake.
RemediationofthelakerequiresastrategytoremovethealgalbiomassPeffectively.
AcknowledgementsThisresearchwasjointlysupportedbytheNationalNaturalScienceFoundationofChina(41703115,41521003,41630645,41807372)andPostdoctoralScienceFoundationofChina(2017M610967).
OpenAccessThisarticleisdistributedunderthetermsoftheCrea-tiveCommonsAttribution4.
0InternationalLicense(http://creativecommons.
org/licenses/by/4.
0/),whichpermitsunrestricteduse,distribu-tion,andreproductioninanymedium,providedyougiveappropriatecredittotheoriginalauthor(s)andthesource,providealinktotheCreativeCommonslicense,andindicateifchangesweremade.
ReferencesAbdiD,Cade-MenunBJ,ZiadiN,ParentLE(2014)Long-termimpactoftillagepracticesandPfertilizationonsoilPformsasdeter-minedby31P-NMRspectroscopy.
JEnvironQual43:1432–1441BaiX,SunJ,ZhouY,GuL,ZhaoH,WangJ(2017)Variationsofdifferentdissolvedandparticulatephosphorusclassesduringanalgaebloominaeutrophiclakeby31PNMRspectroscopy.
Che-mosphere169:577–585BaldwinDS(2013)Organicphosphorusintheaquaticenvironment.
EnvironChem10:439–454BedrockCN,CheshireMV,ChudekJA,FraserAR,GoodmanBA,ShandCA(1995)EffectofpHonprecipitationofhumicacidfrompeatandmineralsoilsonthedistributionofphosphorusformsinhumicandfulvicacidfractions.
CommunSoilSciPlantAnal26:1411–1425BellDW,PellechiaP,ChambersLR,LongoAF,McCabeKM,IngallED,Benitez-NelsonCR(2017)Isolationandmolecularcharac-terizationofdissolvedorganicphosphorususingelectrodialy-sis-reverseosmosisandsolution31P-NMR.
LimnolOceanogr-Methods.
https://doi.
org/10.
1002/lom3.
10171BusatoJG,CanellasLP,RumjanekVM,VellosoACX(2005)Phos-phorusinaninceptisolunderlong-termsugarcane:II.
HumicacidanalysisbyNMRP-31.
RevBrasCiencSolo29:945–953Cade-MenunBJ(2015)Improvedpeakidentificationin31P-NMRspectraofenvironmentalsampleswithastandardizedmethodandpeaklibrary.
Geoderma257:102–114Cade-MenunBJ,LiuCW(2014)Solutionphosphorus-31nuclearmagneticresonancespectroscopyofsoilsfrom2005to2013:areviewofsamplepreparationandexperimentalparameters.
SoilSciSocAmJ78:19–37Cade-MenunBJ,PrestonCM(1996)Acomparisonofsoilextractionproceduresfor31PNMRspectroscopy.
SoilSci161:770–785ChenY,LiuQ(2014)Onthehorizontaldistributionofalgal-bloominChaohuLakeanditsformationprocess.
ActaMechSinca30:656–666DooletteA,SmernikR,DoughertyW(2009)Spikingimprovedsolu-tionphosphorus-31nuclearmagneticresonanceidentificationofsoilphosphoruscompounds.
SoilSciSocAmJ73:919–927FengW,ZhuY,WuF,MengW,GiesyJP,HeZ,SongL,FanM(2016a)Characterizationofphosphorusformsinlakemacro-phytesandalgaebysolution31Pnuclearmagneticresonancespectroscopy.
EnvironSciPollutRes23:7288–7297FengW,ZhuY,WuF,HeZ,ZhangC,GiesyJP(2016b)Formsandlabilityofphosphorusinalgaeandaquaticmacrophytescharacterizedbysolution31PNMRcoupledwithenzymatichydrolysis.
SciRep6:37164FengW,WuF,HeZ,SongF,ZhuY,GiesyJP,WangY,QinN,ZhangC,ChenH,SunF(2018)Simulatedbioavailabilityofphosphorusfromaquaticmacrophytesandphytoplanktonbyaqueoussuspensionandincubationwithalkalinephosphatase.
SciTotalEnviron616:1431–1439GilesCD,Cade-MenunBJ(2014)Appliedmanureandnutrientchemistryforsustainableagricultureandenvironment.
In:ZhangH(ed)Phytateinanimalmanureandsoils:abundance,cyclingandbioavailability.
Springer,TheNetherland,pp163–190GilesCD,LeeLG,Cade-MenunBJ,HillJE,IslesPD,SchrothAW,DruschelGK(2015)CharacterizationoforganicphosphorusformandbioavailabilityinlakesedimentsusingPnuclearmagneticresonanceandenzymatichydrolysis.
JEnvironQual44:1–13HeZ,HoneycuttCW(2005)Amodifiedmolybdatebluemethodfororthophosphatedeterminationsuitableforinvestigatingenzymatichydrolysisoforganicphosphates.
CommunSoilSciPlantAnal36:1373–1383HeZ,Cade-MenunBJ,ToorGS,SimJT(2007)Comparisonofphos-phorusformsinwetanddriedanimalmanuresbysolutionphos-phorus-31nuclearmagneticresonancespectroscopyandenzy-matichydrolysis.
JEnvironQual36:1086–1095HeZ,HoneycuttCW,Cade-MenunBJ,SenwoZN,TazisongIA(2008)Phosphorusinpoultrylitterandsoil:enzymaticandnuclearmag-neticresonancecharacterization.
SoilSciSocAmJ72:1425–1433HeZ,HoneycuttCW,GriffinTS,Cade-MenunBJ,PellechiaPJ,DouZ(2009)Phosphorusformsinconventionalandorganicdairy171Limnology(2019)20:163–171manureidentifiedbysolutionandsolidstateP-31NMRspectros-copy.
JEnvironQual38:1909–1918HeZ,OlkDC,Cade-MenunBJ(2011)Formsandlabilityofphospho-rusinhumicacidfractionsofHordsiltloamsoil.
SoilSciSocAmJ75:1712–1722JaroschKA,DooletteAL,SmernikRJ,TamburiniF,FrossardE,Büne-mannEK(2015)CharacterisationofsoilorganicphosphorusinNaOH–EDTAextracts:acomparisonof31PNMRspectroscopyandenzymeadditionassays.
SoilBiolBiochem91:298–309KagalouaI,PapastergiadoubE,LeonardasaI(2008)Long-termchangesintheeutrophicationprocessinashallowMediterraneanlakeecosystemofWGreece:responseafterthereductionofexter-nalload.
JEnvironManag87:497–506KarlDM(2014)Microbiallymediatedtransformationsofphosphorusinthesea:newviewsofanoldcycle.
AnnRevMarSci6:279–337LehmanPW,KurobeT,LesmeisterS,BaxaD,TungA,TehSJ(2017)Impactsofthe2014severedroughtontheMicrocystisbloominSanFranciscoestuary.
HarmfulAlgae63:94–108LiM,WuY,YuZ,ShengG,YuH(2009)Enhancednitrogenandphos-phorusremovalfromeutrophiclakewaterbyIpomoeaaquaticawithlow-energyionimplantation.
WaterRes43:1247–1256LiJ,CuiK,LuW,ChengY,JiangY(2015)Communitydynamicsofspring-summerplanktoninLakeChaohu.
ActaHydrobiolSinica39:185–194(inChinese)LiuS,ZhuY,MengW,HeZ,FengW,ZhangC,GiesyJP(2016)Char-acteristicsanddegradationofcarbonandphosphorusfromaquaticmacrophytesinlakes:insightsfromsolid-state13CNMRandsolu-tion31PNMRspectroscopy.
SciTotalEnviron543:746–756LiuS,ZhuY,WuF,MengW,WangH,HeZ,GuoW,SongF,GiesyJP(2017)Usingsolid13CNMRcoupledwithsolution31PNMRspectroscopytoinvestigatemolecularspeciesandlabilityoforganiccarbonandphosphorusfromaquaticplantsinTaiLake.
ChinaEnvironSciPollutRes24:1880–1889MahieuN,OlkDC,RandallEW(2000)Analysisofphosphorusintwohumicacidfractionsofintensivelycroppedlowlandricesoilsby31PNMR.
EurJSoilSci51:391–402McDowellRW,HillSJ(2015)Speciationanddistributionoforganicphosphorusinriversediments:anationalsurvey.
JSoilSediment15:2369–2379Pernet-CoundrierB,QiWX,LiuHJ,NüllerB,BergM(2012)Sourcesandpathwayofnutrientsinthesemi-aridregionofBeijing–Tian-jin.
ChinaEnvironSciTechnol46:5294–5301QuX,XieL,LinY,BaiY,ZhuY,XieF,GeisyJP,WuF(2013)Quan-titativeandqualitativecharacteristicsofdissolvedorganicmatterfromeightdominantaquaticmacrophytesinLakeDianchi.
ChinaEnvironSciPollutRes20:7413–7423ReitzelK,AhlgrenJ,GogollA(2006)Effectsofaluminumtreatmentonphosphorus,carbon,andnitrogendistributioninlakesediment:a31PNMRstudy.
WaterRes40:647–654RubanV,López-SánchezJF,PardoP(2001)Harmonizedprotocolandcertifiedreferencematerialforthedeterminationofextractablecontentsofphosphorusinfreshwatersediments—asynthesisofrecentworks.
FresenJAnalChem370:224–228SchneiderK,ReslP,SpribilleT(2016)Escapefromthecrypticspeciestrap:lichenevolutiononbothsidesofacyanobacterialacquisitionevent.
MolEcol25:3453–3468SmernikRJ,DoughertyWJ(2007)Identificationofphytateinphos-phorus-31nuclearmagneticresonancespectra:theneedforspik-ing.
SoilSciSocAmJ71:1045–1050SrensenDR,NielsenUG,SkouEM(2014)Solidstate31PMASNMRspectroscopyandconductivitymeasurementsonNbOPO4andH3PO4compositematerials.
JSolidStateChem219:80–86SuzumuraM(2005)Phospholipidsinmarineenvironments:areview.
Talanta66:422–434TangJ,ShiT,WuX,CaoH,LiX,HuaR,TangF,YueY(2015)TheoccurrenceanddistributionofantibioticsinLakeChaohu,China:seasonalvariation,potentialsourceandriskassessment.
Chem-osphere122:154–161TurnerBL,RichardsonAE(2004)Identificationof-inositolphosphatesinsoilbysolutionphosphorus-31nuclearmagneticresonancespectroscopy.
SoilSciSocAmJ68:802–808TurnerBL,MahieuN,CondronLM(2003)Phosphorus-31nuclearmagneticresonancespectralassignmentsofphosphoruscom-poundsinsoilNaOH–EDTAextracts.
SoilSciSocAmJ67:497–510TurnerBL,Cade-MenunBJ,CondronLM,NewmanS(2005)Extrac-tionofsoilorganicphosphorus.
Talanta66:294–306TurnerBL,CheesmanAW,GodageHY,RileyAM,PotterBVL(2012)Determinationofneo-andd-chiro-inositolhexakisphosphateinsoilsbysolution31PNMRspectroscopy.
EnvironSciTechnol46:4994–5002VincentAG,VestergrenJ,GrbnerG,PerssonP,SchleucherJ,BieslerR(2013)Soilorganicphosphorustransformationsinaborealfor-estchronosequence.
PlantSoil367:149–162WangJ,PantHK(2010)Enzymatichydrolysisoforganicphosphorusinriverbedsediments.
EcoEng36:963–968WangX,XiB,HuoS,DengL,PanH,XiaX,ZhangJ,RenY,LiuH(2013)PolybrominateddiphenylethersoccurrenceinmajorinflowingriversofLakeChahu(China):characteristics,potentialsourcesandinputstolake.
Chemosphere93:1624–1631XuM,CaoH,XieP,DengD,FengW,XuJ(2005)Thetemporalandspatialdistribution,compositionandabundanceofprotozoainChaohuLake,China:relationshipwitheutrophication.
EuropJProtistol41:183–192YangL,HanX,SunP,YanW,LiY(2011)CanonicalcorrespondenceanalysisofalgaecommunityanditsenvironmentalfactorsinLakeChaohu.
ChinaJAgro-EnvironSci30:952–958YoungEO,RossDS,Cade-MenunBJ,LiuCW(2013)Phosphorusspeciationinripariansoils:aphosphorus-31nuclearmagneticresonancespectroscopyandenzymehydrolysisstudy.
SoilSciSocAmJ77:1636–1647ZanF,HuoS,XiB,LiQ,LiaoH,ZhangJ(2010)Phosphorusdistribu-tioninthesedimentsofashalloweutrophiclake.
LakeChaohu,China.
EnvironEarthSci62:1643–1653ZhangWQ,XingBS(2013)DetectionofphosphorusspeciesinthesedimentsofChaohuLakeby31Pnuclearmagneticresonancespectroscopy(31P-NMR).
ActaSciCircum33:1967–1973(inChinese)ZhangWQ,ZhuXL,JinX,MengX,TangWZ,ShanBQ(2017)Evi-dencefororganicphosphorusactivationandtransformationatthesediment-waterinterfaceduringplantdebrisdecomposition.
SciTotalEnviron583:458–465ZhongW,ZhangZ,LuoY(2012)Biogasproductivitybyco-digestingTaihubluealgaewithcornstrawasanexternalcarbonsource.
BioresourceTechnol114:281–286ZhuG,QinB,ZhangL,LuoL(2006)Geochemicalformsofphos-phorusinsedimentsofthreelarge,shallowlakesofChina.
Pedo-sphere16:726–734ZhuY,WuF,HeZ,GuoJ,QuX,XieF,GiesyJP,LiaoH,GuoF(2013)Characterizationoforganicphosphorusinlakesedimentsbysequentialfractionationandenzymatichydrolysis.
EnvironSciTechnol47:7679–7687ZhuY,WuF,HeZ(2015)Bioavailabilityandpreservationoforganicphosphorusinfreshwatersedimentsanditsroleinlakeeutrophi-cation.
In:HeZ,WuF(eds)Labileorganicmatter—chemicalcompositions,function,andsignificanceinsoilandtheenviron-ment.
SSSA,Madison,pp275–294ZhuY,WuF,FengW,LiuS,GiesyJP(2016)Interactionofalkalinephosphatasewithmineralsandsediments:activities,kineticsandhydrolysisoforganicphosphorus.
ColloidssurfA:PhysicochemEngAspects495:46–53
Bluehost美国虚拟主机2.95美元/月,十八周年庆年付赠送顶级域名和SSL证书
Bluehost怎么样,Bluehost好不好,Bluehost成立十八周年全场虚拟主机优惠促销活动开始,购买12个月赠送主流域名和SSL证书,Bluehost是老牌虚拟主机商家了,有需要虚拟主机的朋友赶紧入手吧,活动时间:美国MST时间7月6日中午12:00到8月13日晚上11:59。Bluehost成立于2003年,主营WordPress托管、虚拟主机、VPS主机、专用服务器业务。Blueho...
RAKsmart:美国洛杉矶独服,E3处理器/16G/1TB,$76.77/月;美国/香港/日本/韩国站群服务器,自带5+253个IPv4
RAKsmart怎么样?RAKsmart机房即日起开始针对洛杉矶机房的独立服务器进行特别促销活动:低至$76.77/月,最低100Mbps带宽,最高10Gbps带宽,优化线路,不限制流量,具体包括有:常规服务器、站群服务器、10G大带宽服务器、整机机柜托管。活动截止6月30日结束。RAKsmart,美国华人老牌机房,专注于圣何塞服务器,有VPS、独立服务器等。支持PayPal、支付宝付款。点击直达...
Spinservers:美国圣何塞机房少量补货/双E5/64GB DDR4/2TB SSD/10Gbps端口月流量10TB/$111/月
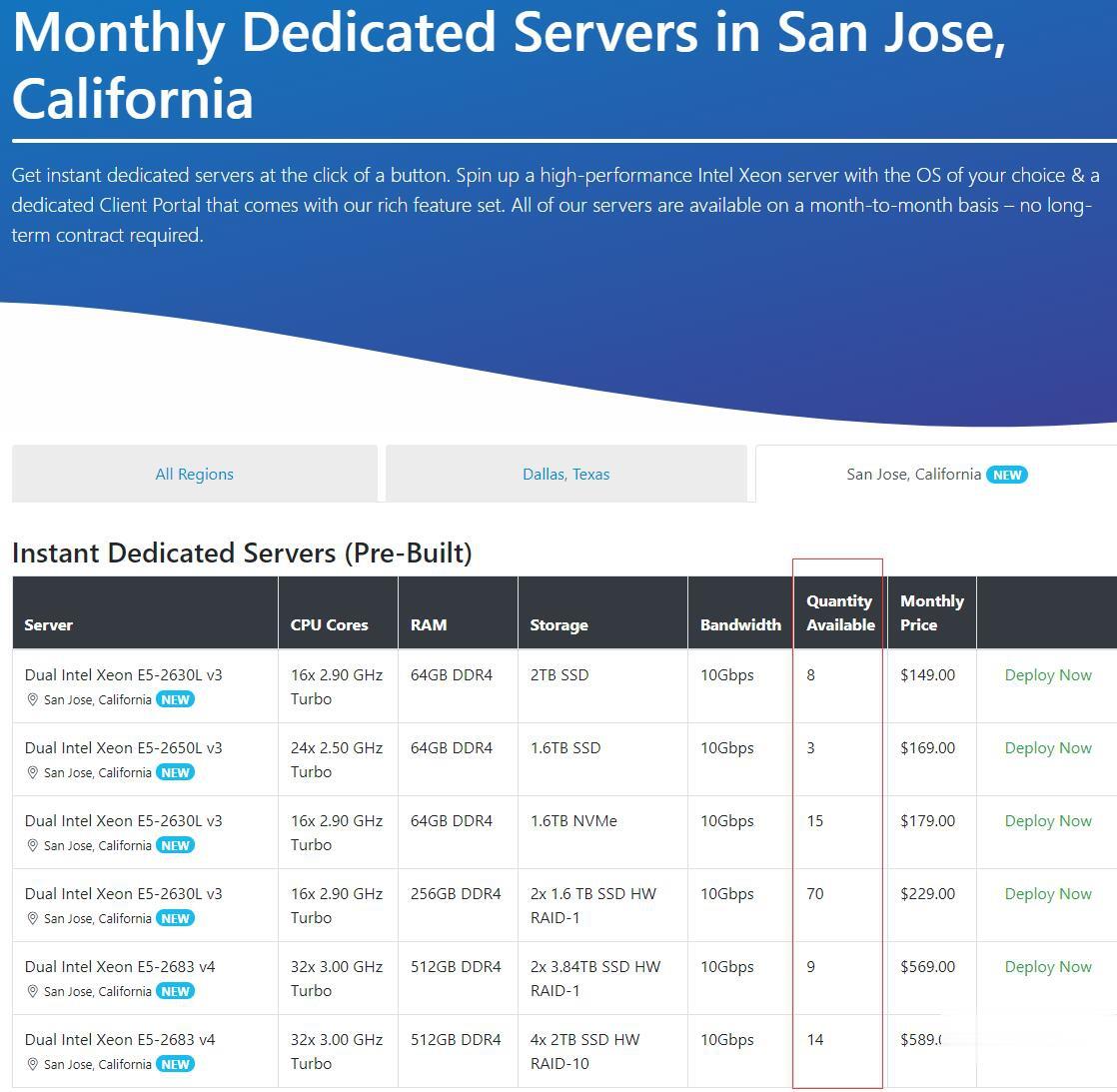
Chia矿机,Spinservers怎么样?Spinservers好不好,Spinservers大硬盘服务器。Spinservers刚刚在美国圣何塞机房补货120台独立服务器,CPU都是双E5系列,64-512GB DDR4内存,超大SSD或NVMe存储,数量有限,机器都是预部署好的,下单即可上架,无需人工干预,有需要的朋友抓紧下单哦。Spinservers是Majestic Hosting So...
网易yeah为你推荐
-
重庆大学网上服务大厅用新iphone也将禁售iPhone8plus在2020年还会有货吗建企业网站建立一个企业网站要多少费用cisco2960配置Cisco2960是二层交换机,怎么可以进入配置界面进行配置。不是说二层交换机不需要配置吗?字节跳动回应TikTok易主一部电影讲一个小伙子去继承遗产结果是一批雪橇狗男主吹口哨声明不是雪地狂奔asp.net网页制作如何用ASP.NET做网站?客服电话各银行的客服电话是多少?抢米网会知道怎样抢小米的请进来说一下。银花珠树晓来看晚来天欲雪,能饮一杯无。相似的句子12306.com注册12306邮箱地址怎么写