step66smsm.com
66smsm.com 时间:2021-03-19 阅读:()
RapidCommunicationStructuralbrainvariationandgeneralintelligenceRichardJ.
Haier,a,*RexE.
Jung,bRonaldA.
Yeo,cKevinHead,aandMichaelT.
AlkiredaDepartmentofPediatrics,UniversityofCalifornia,Irvine,CA92697-5000,USAbDepartmentofNeurology,andtheMINDInstitute,UniversityofNewMexico,Albuquerque,NM87131,USAcDepartmentofPsychology,UniversityofNewMexico,Albuquerque,NM87131,USAdDepartmentofAnesthesiology,UniversityofCaliforniaIrvineMedicalCenter,Orange,CA92868-1350,USAReceived10March2004;revised14April2004;accepted22April2004Availableonline15July2004Totalbrainvolumeaccountsforabout16%ofthevarianceingeneralintelligencescores(IQ),buthowvolumesofspecificregions-of-interest(ROIs)relatetoIQisnotknown.
Weusedvoxel-basedmorphometry(VBM)intwoindependentsamplestoidentifysubstantialgraymatter(GM)correlatesofIQ.
Basedonstatisticalconjunctionofbothsamples(N=47;P<0.
05correctedformultiplecomparisons),moregraymatterisassociatedwithhigherIQindiscreteBrodmannareas(BA)includingfrontal(BA10,46,9),temporal(BA21,37,22,42),parietal(BA43and3),andoccipital(BA19)lobesandnearBA39forwhitematter(WM).
Theseresultsunderscorethedistributedneuralbasisofintelligenceandsuggestadevelopmentalcourseforvolume–IQrelationshipsinadulthood.
D2004ElsevierInc.
Allrightsreserved.
Keywords:IQ;Brainvolume;MorphometryIntroductionCorrelationsbetweenregionalbrainfunctionandperformanceonmentaltestsassociatedwithageneralintelligencefactor(g)asdefinedoriginallybySpearman(1904)havebeendemonstratedmanytimesinnormalsubjects(Duncanetal.
,2000;Grayetal.
,2003;Haieretal.
,1988;HaierandBenbow,1995;Parksetal.
,1988;Prabhakaranetal.
,1997).
Mostofthesestudiesshowthatgoodtestperformancerecruitsareasdistributedthroughoutthebrain,althoughacasehasbeenmadethatactivationwithinareasofthefrontallobesistheprimarysourceofdifferencesing-loadedtestperformance(Duncanetal.
,2000).
Thereisevidencethatdeactivationwithinsomebrainareas,includingfrontallobes,isassociatedwithbettermentaltaskperformance(Haieretal.
,1988,1992a;Parksetal.
,1988),especiallyinsubjectswithhigherintelligencetestscores(Haieretal.
,1992b).
Evenwhenapassivetaskwithnoinherentproblemsolvingisused,subjectswithhigherintelligencescoresshowmoreactivationinposteriorinformationprocessingareasthansubjectswithlowerscores(Boivinetal.
,1992;Haieretal.
,2003b).
Functionalbrainimagingstudiesalwaysmustbeinterpretedtotakeaccountofthespecifictaskdemandsofthementaltaskusedduringtheimagingprotocol.
Thismakesinconsistenciesamongstudyresultsdifficulttoreconcilegiventhewidevarietyoftasksused.
Totheextentthatindividualdifferencesingeneralintelli-gencehaveastructuralcomponent,examiningstructuralcorrelatesofintelligencewouldeliminateanytask-relatedinfluencesfromconsideration.
Forthisreason,structuralimagingofregionalgrayandwhitemattervolumeswouldprovideuniqueinformationaboutthedistributionofbrainareasrelatedtogeneralintelligence.
Forexample,totalbrainvolumeassessedbyMRIinmanystudieshasbeenshowntocorrelateaboutr=0.
40,withintelli-gencescoresandtotalgrayandwhitemattervolumesalsoshowsmallcorrelationswithIQ(Gignacetal.
,2003),butattemptstorelatevolumeofspecificbrainareastotestscoreshavebeenmostlyunsuccessful(Flashmanetal.
,1997;MacLullichetal.
,2002).
Untilnow,suchattemptshaveusedvariousregion-of-interest(ROI)methodsthataredifficulttoreliablyapplytomanybraingyriwhenoutlinedbyhandandoftendonotconformwelltotheextensiveindividualdifferencesamongsubjectsinbrainsizeandmorphologywhenappliedstereotactically.
Arecentmethod-ologicaladvanceisoptimizedvoxel-basedmorphometry(VBM),whichusesalgorithmstosegmentgraymatter(GM)andwhitematter(WM)fromstructuralMRIs(AshburnerandFriston,2000;Goodetal.
,2001).
VBMhasbeenvalidatedextensively(Ash-burnerandFriston,2001;Goodetal.
,2002)andithasbeenused,forexample,tocharacterizegrayandwhitemattervolumechangesinaging(Goodetal.
,2001),dementia(Goodetal.
,2002),andDownsyndrome(Whiteetal.
,2003).
MaterialsandmethodsSubjectsWetestedtwosamplesandusedastatisticalconjunctionapproach(PriceandFriston,1997)toshowwherecorrelationsbetweenIQandgrayorwhitematterwerecommontobothsamples.
Thefirstsamplewas23normalvolunteers(14womenand9men;1053-8119/$-seefrontmatterD2004ElsevierInc.
Allrightsreserved.
doi:10.
1016/j.
neuroimage.
2004.
04.
025*Correspondingauthor.
DepartmentofPediatrics,UniversityofCaliforniaIrvine,MedicalScienceI,B140,Irvine,CA92697.
Fax:+1-949-824-9059.
E-mailaddress:rjhaier@uci.
edu(R.
J.
Haier).
AvailableonlineonScienceDirect(www.
sciencedirect.
com.
)www.
elsevier.
com/locate/ynimgNeuroImage23(2004)425–433meanage=27,SD=5.
9,range=18–37)recruitedfromtheUniversityofNewMexico(UNM).
Sample1MRIswereobtainedwitha1.
5-Tscanner,headcoil,andsoftware(Signa5.
4;GeneralElectricMedicalSystems,Waukesha,WI).
AT1sagittallocalizersequence(TE=6.
9ms,TR=200ms,FOV=2424cm2,fiveslices,thickness=5mm,spacing=2.
5mm,matrix=256128)wasacquired,followedbyaT1-weightedaxialseries(fastRFspoiledgradient-recalled,TE=6.
9ms,TR=17.
7ms,flipangle=25j,matrix=256192,120slices,thickness=1.
5mm)togivefullbraincoverage.
Thesecondsamplewas24normalvolunteers(13menand11women,meanage=59,SD=15.
9,range37–84)recruitedattheUniversityofCalifornia(UCI),Irvine,asmiddle-agedandoldernormalcontrolsforanimagingstudyofdementiainDownsyndromeandAlzheimer'sdisease(Haieretal.
,2003a).
Sample2MRIswereobtainedwitha1.
5-TclinicalPhillipsEclipsescanner(PhilipsMedicalSystems,N.
A.
,Bothell,WA).
WeusedT1-weighted,volumetricSPGRMRIscans(FOV=24cm,flipangle=40,TR=24,TE=5).
Theimagesconsistedof120contiguous1.
2-mmthickaxialslices,eachwithanin-planeimagematrixof256256imageelements.
Allimagesinbothsampleswerevisuallyinspectedtoensureimagequality.
IntelligencetestingToassessgeneralintelligence,subjectsweretestedwiththeWechslerAdultIntelligenceScale(WAIS).
TheWAISbattery(Wechsler,1981)consistsof11diversesubtests,whichtapavarietyofverbalandnonverbalmentalabilitiesthatcontributetogeneralintelligence.
TheWAISFullScaleIQ(FSIQ)scoreisbasedonperformanceofall11subtests(accordingtoagebasednorms).
Factoranalyticstudies(Jensen,1980)showthateachsubtestloadsonthegfactorandtheFSIQscoreloadsthehighest(about0.
90or81%ofthevarianceing).
Forthisreason,FSIQisconsideredoneofthebestindexesofindividualdifferencesingeneralintelligence.
Forsample1(UMN),theFSIQwas116(SD=14.
7)andtherangewas90–155.
Forsample2(UCI),themeanFSIQwas116(SD=14.
2)andtherangewas90–142.
Voxel-basedmorphometryWeappliedVBMtoidentifybrainareaswhereGMandWMvolumesarecorrelatedtoFSIQtotestwhetheranysuchareasareclusteredinfrontallobesordistributedthroughoutthebrain.
WeusedStatisticalParametricMappingsoftware(SPM2;TheWellcomeDepartmentofImagingNeuroscience,UniversityCollegeLondon)tocreateastudy-specifictemplateandthenappliedtheoptimizedVBMprotocoltoeachsampleseparatelyusingthemethodsofAshburnerandFriston(2000)andGoodetal.
(2001).
Topreservetheamountoftissueinanygivenanatomicalregionafterspatialnormalization,theoptimalGMandWMpartitionsweremultipliedbytheJacobiandetermi-nantsoftheirrespectivespatialtransformationmatrix.
ThereasonforperformingthismodulationstepissothatthefinalVBMstatisticswillreflectlocaldeviationsintheabsoluteamount(volume)oftissueindifferentregionsofthebrain(AshburnerandFriston,2000).
ThemodulatedGMandWMpartitionswerethensmoothedwitha12-mmFWHMisotropicGaussiankerneltoaccountforslightmisalignmentsofhomol-ogousanatomicalstructuresandtoensurestatisticalvalidityunderparametricassumptions.
StatisticalconjunctionapproachWespecificallytestedwhetherregionalgrayandwhitemattervolumeswerecorrelatedwithFSIQscorestreatinganyeffectsofage,sex,andhandedness(threecasesintheUNMsamplewerelefthanded)asnuisancevariablesintheSPM2designmatrix.
Aftercomputingthecorrelationanalysesforeachsampleseparately,weFig.
1.
CorrelationsbetweengrayandwhitematterandFSIQconjunctedacrossbothsamples.
Basedonstatisticalconjunction,correlationsbetweengraymatterandFSIQareshownintheleftcolumnongraymattertemplates;whitemattercorrelationstoFSIQareshownintherightcolumnonawhitemattertemplate.
Toprowshowsfrontalview,secondrowshowsmedialsagittalview,nextrowsshowleftandrightlateralviews,andbottomrowshowsoccipitalview.
Anatomicallocations,atlascoordinates,andclustersizesarelistedinTable1.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433426Table1LocalizationofconjunctioncorrelationsfromUNMandUCIbetweengrayandwhitematterwithFSIQ(PV0.
001)Brainarea(and/ornearestgraymatter)Clustersizexyztr2GraymattercorrelationswithFSIQRightmedialfrontalgyrus,BA10*684626664.
650.
35Rightmedialfrontalgyrus,BA10*863174.
380.
32Rightsuperiorfrontalgyrus,BA9*456314.
290.
31Leftmiddlefrontalgyrus,BA46*34645039174.
320.
31Leftinferiorfrontalgyrus,BA45*513154.
130.
29Leftmiddlefrontalgyrus,BA46*5035243.
920.
27Leftmiddletemporalgyrus,BA21*692664543.
930.
27Rightfrontalprecentralgyrus,BA9*7053723343.
860.
27Rightmiddlefrontalgyrus,BA9*4932323.
220.
20Righttemporalinferiorgyrus,BA37*22945560103.
840.
26Righttemporalmiddlegyrus,BA37*575183.
650.
25Righttemporalmiddlegyrus,BA21*614833.
510.
23Leftmiddlefrontalgyrus,BA10*524405633.
750.
26Leftsuperiorfrontalgyrus,BA10*3651233.
650.
25Leftmiddlefrontalgyrus,BA10*415393.
510.
23Leftmiddleoccipitalgyrus,BA19*345546853.
660.
25Lefttemporalsuperiorgyrus,BA22*4266348193.
610.
24Leftparietalsupramarginalgyrus,BA40*6645263.
420.
22Lefttemporaltransversegyrus,BA42*218629123.
470.
23Rightparietalpostcentralgyrus,BA43*3856417153.
460.
23Rightparietalpostcentralgyrus,BA3*6115233.
360.
22Righttemporalmiddlegyrus,BA21*456535103.
300.
21Leftfrontalsuperiorgyrus,BA8*134123503.
280.
21Rightfrontalinferiorgyrus,BA9*20587303.
280.
21WhitemattercorrelationswithFSIQRighttemporalmiddlegyrus,BA39*4457515453.
480.
23Righttemporalfusiformgyrus,BA375037103.
240.
20Rightmiddletemporalgyrus,BA21553332.
510.
13Lefttemporalmiddlegyrus,BA204325524093.
380.
22Lefttemporalinferiorgyrus,BA19445313.
140.
19Lefttemporalmiddlegyrus,BA37525322.
700.
15Leftfrontalsuperiorgyrus,BA101839186014.
550.
34Leftfrontalmedialgyrus,BA1076383.
920.
27Leftfrontalmedialgyrus,BA10862213.
470.
23Leftfrontalinferiorgyrus,BA478132313132.
980.
18Rightfrontalmiddlegyrus,BA102126295982.
930.
17Rightfrontalsuperiorgyrus,BA10216262.
850.
17Rightfrontalmedialgyrus,BA102444102.
360.
12Leftparietalprecuneus,BA726752360302.
790.
16Leftparietalprecuneus,BA71552242.
370.
12Rightparietalprecuneus,BA3112881858352.
790.
16Rightlimbiccingulategyrus,BA2418191315322.
750.
16Rightlimbiccingulategyrus,BA32139362.
300.
11Rightfrontalmedialgyrus,BA81131392.
240.
11Leftfrontalsuperiorgyrus,BA95821052352.
670.
15Leftfrontalsuperiorgyrus,BA9958322.
170.
10Rightlimbic,parahippocampus,BA357261726212.
550.
14Rightlimbic,parahippocampus,BA351930142.
360.
12Leftlimbic,parahippocampus,BA3515801428192.
460.
13Leftbrainstem,pons431222.
270.
11Leftbrainstem,midbrainsubstanianigra1127112.
210.
11Rightfrontalsuperiorgyrus,BA113242949152.
410.
12Rightfrontalsuperiorgyrus,BA112456152.
110.
10Leftsublobar,insula,BA13245438122.
400.
12Rightcerebellum,anteriorlobe4063046302.
360.
12Rightfrontalsuperiorgyrus,BA63135542.
200.
11Leftfrontalsuperiorgyrus,BA111402047172.
200.
11Leftfrontalprecentralgyrus,BA660491112.
180.
10Righttemporal,subgyral,BA20894216172.
170.
10Leftfrontalsuperiorgyrus,BA658920532.
140.
10x,y,andzcoordinatesareinTalairachatlasspace.
*P<0.
05correctedformultiplecomparisons.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433427usedtheconjunctionapproach(PriceandFriston,1997)toshowwheregraymatter(andwhitematter)correlationsoverlappedfortheUNMandtheUCIsamples(i.
e.
,voxelswithcorrelationsincommonforbothsamples).
Thisconjunctionapproachminimizespotentialproblemsassociatedwithcombiningdatafromdifferentscanners.
Theconjunctionanalysisalsohastheadvantageofmaximizingstatisticalpowerbecauseall47subjectsareusedintheanalysisanditisequivalenttoafixedfactormodelinSPM.
FindingsareconsideredsignificantatP<0.
05correctedformultiplecomparisons;findingsatPV0.
001uncorrectedalsoareshownforhypothesisgeneration.
R2estimatesweredeterminedusingtheformula:R2=t2/(df+t2)wheredf=41(fromtheSPM2conjunctiondesignmatrix).
Locationsofsignificantclusters(cent-roids)areconvertedfromMontrealNeurologicalInstitute(MNI)toTalairachatlas(TalairachandTournoux,1988)coordinatesandreportedasclosestBrodmannarea(BA)wherepossible.
Onlyclustersofatleast10voxelsarereported.
ResultsTheconjunctionresults(N=47;Fig.
1)showedrobustpositivecorrelations(P<0.
05,correctedformultiplecomparisons)betweenFSIQandgraymattervolumesinBrodmannareas(BA)10,46,and9infrontallobes;BA21,37,22,and42intemporallobes;BA43and3inparietallobes;andBA19intheoccipitallobe.
ThesizeandFig.
2.
CorrelationsbetweengrayandwhitematterandFSIQforeachsample.
Basedonseparateanalysesforeachsample,graymattercorrelationsareshownintheleftcolumns(UNMandUCI,respectively);whitemattercorrelationsareshownintherightcolumns.
Toprowshowsfrontalview,secondrowshowsmedialsagittalview,nextrowsshowleftandrightlateralviews,andbottomrowshowsoccipitalview.
Anatomicallocations,atlascoordinates,andclustersizesarelistedinTables2(UNMdata)and3(UCIdata).
R.
J.
Haieretal.
/NeuroImage23(2004)425–433428locationsoftheseareasareshowninTable1.
Similarbutlessrobustcorrelationswithwhitematterareas(PV0.
001,uncorrected)werealsofound(Table1).
Notably,24graymatterregionsweresigni-ficantatP<0.
05(corrected),yetonlyonewhitematterregionwassignificantatthislevel.
EstimatesofFSIQvariabilityaccountedforbythehighestcorrelationforanindividualvoxelwithinthesignificantgrayandwhitematterareas(R2)areashighas73%(Table1).
Therewerenosignificantnegativecorrelations.
Table2LocalizationofcorrelationsintheUNMsamplebetweengrayandwhitematterwithFSIQ(PV0.
001)Brainarea(and/ornearestgraymatter)Clustersizexyztr2GraymattercorrelationswithFSIQLefttemporalfusiformgyrus,BA37*3984415296.
930.
73Lefttemporalfusiformgyrus,BA364439274.
170.
49Lefttemporalfusiformgyrus,BA363943234.
090.
48Leftfrontalsuperiorgyrus,BA1018,470760256.
280.
69Rightfrontalsuperiorgyrus,BA10763186.
030.
67Rightfrontalsuperiorgyrus,BA8943545.
510.
63Leftfrontalsuperiorgyrus,BA61086017625.
990.
67Righttemporalfusiformgyrus,BA3610,1874436305.
940.
66Righttemporalfusiformgyrus,BA205140275.
290.
61Righttemporalmiddlegyrus,BA37576215.
180.
60Rightlimbiclobe,anteriorcingulate,BA321756143075.
360.
61Rightlimbiclobe,anteriorcingulate,BA321731174.
420.
52Rightfrontalinferiorgyrus,BA479623113205.
230.
60Leftparietalangulargyrus,BA392513058344.
810.
56Leftlimbiclobe,anteriorcingulate,BA328301738164.
780.
56Leftfrontalmiddlegyrus,BA10602405874.
710.
55Leftfrontalmiddlegyrus,BA103952133.
930.
46Rightfrontalsuperiorgyrus,BA111213046224.
660.
55Leftsublobar,insula,BA13151743344.
630.
54Leftsublobar,insula,BA13371374.
000.
47Leftoccipitalmiddlegyrus,BA19283616594.
480.
53Leftlimbiclobe,cingulategyrus,BA23338816304.
210.
50Leftlimbiclobe,cingulategyrus,BA2496323.
730.
44Rightmedialfrontalgyrus,BA10275135144.
160.
49Leftmiddletemporalgyrus,BA22597514714.
160.
49Leftfrontalinferiorgyrus,BA476252924194.
150.
49Leftfrontalinferiorgyrus,BA473828164.
100.
48Leftfrontalinferiorgyrus,BA472516183.
910.
46Lefttemporalfusiformgyrus,BA201136011284.
030.
47Rightfrontalinferiorgyrus,BA92725421243.
970.
47Leftfrontalmiddlegyrus,BA461854841203.
910.
46Leftfrontalmiddlegyrus,BA8174224493.
890.
46Rightfrontalmiddlegyrus,BA8493830493.
890.
46Leftlimbiclobe,anteriorcingulate,BA10691232113.
850.
45Rightfrontalmiddlegyrus,BA10334240183.
830.
45Rightfrontalmiddlegyrus,BA9944121343.
790.
44Rightlimbiclobe,parahippocampus,BA1929344503.
790.
44Leftfrontalmedialgyrus,BA32201414443.
750.
44Leftinferiorfrontalgyrus,BA4735542133.
740.
44Lefttemporalsuperiorgyrus,BA38203613203.
710.
43WhitemattercorrelationswithFSIQLeftfrontalsuperiorgyrus,BA9690850215.
870.
66Leftfrontalmedialgyrus,BA9844293.
840.
45Rightfrontalmedialgyrus,BA102234845145.
190.
60Rightfrontalmedialgyrus,BA101847114.
640.
54Rightfrontalsuperior,BA102555134.
070.
48Rightlimbiclobe,parahippocampus,amygdala245287225.
080.
59Rightfrontalsuperiorgyrus,BA84801128485.
010.
58Rightmiddletemporalgyrus,BA21148595244.
950.
58Rightsublobar,insula,BA221138442714.
530.
53Lefttemporalmiddlegyrus,BA202055037104.
320.
51Leftmedialfrontalgyrus,BA10345105754.
140.
49Leftfrontalsuperiorgyrus,BA1076023.
830.
45Rightbrainstem,pons1461421263.
970.
47x,y,andzcoordinatesareinTalairachatlasspace.
*P<0.
05correctedformultiplecomparisons.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433429ThepositivecorrelationsbetweengrayandwhitematterandFSIQforeachsampleseparatelyareshowninFig.
2andinTables2(UNM)and3(UCI);therewerenosignificantnegativecorre-lations.
StatisticalcomparisonsmadedirectlybetweenthetwosamplescouldbedifficulttointerpretbecausedifferentMRIscannerswereusedandtheagesweredifferent.
Anexploratoryanalysisshowedfewareaswithsignificantdifferencesbetweengrayandwhitemattervolumesbetweenthesamples,andnoneoftheseareasoverlappedwithareasidentifiedintheconjunctionanalysis.
AnotherapproachtoassessingthedifferencesbetweenthesamplesisshowninTable4.
ItshowsacomparisonbetweentheTable3LocalizationofcorrelationsintheUCIsamplebetweengrayandwhitematterwithFSIQ(PV0.
001)Brainarea(and/ornearestgraymatter)Clustersizexyztr2GraymattercorrelationswithFSIQLeftfrontalinferiorgyrus,BA45*11,086553017.
060.
70Leftfrontalinferiorgyrus,BA465632105.
500.
59Leftfrontalmiddlegyrus,BA10425535.
350.
58Lefttemporalmiddlegyrus,BA212424644535.
150.
56Leftoccipitalmiddlegyrus,BA19536734.
370.
48Lefttemporalmiddlegyrus,BA21673393.
940.
43Rightfrontalprecentralgyrus,BA951773723345.
130.
56Rightfrontalmiddlegyrus,BA465332214.
330.
47Rightfrontalmiddlegyrus,BA94525334.
310.
47Rightfrontalmedialgyrus,BA10186926665.
030.
55Leftfrontalmedialgyrus,BA10565114.
430.
48Rightfrontalsuperiorgyrus,BA102266114.
270.
46Rightfrontalprecentralgyrus,BA63096607414.
830.
53Rightfrontalprecentralgyrus,BA66510324.
230.
46Rightparietalpostcentralgyrus,BA436413204.
180.
45Leftlingualgyrus4378117714.
720.
51Leftoccipitalfusiformgyrus,BA182187124.
180.
45Leftoccipitalcuneus,BA1809664.
010.
43Rightparietalprecuneus,BA7433961644.
580.
50Rightfrontalinferiorgyrus,BA46296363464.
400.
48Leftcerebellum,posteriorlobe3053276224.
120.
45Leftparietalinferiorlobule,BA404875559394.
100.
44Leftparietalinferiorlobule,BA405450463.
790.
41Rightparietalinferiorlobule,BA401115143504.
070.
44Leftparietalpostcentralgyrus,BA780953704.
060.
44Rightfrontalsuperiorgyrus,BA9269458284.
000.
43Rightfrontalmedialgyrus,BA10462203.
730.
40Leftfrontalmiddlegyrus,BA93045216253.
990.
43Lefttemporalmiddlegyrus,BA21446910173.
920.
42Leftfrontalsuperiorgyrus,BA10623651233.
910.
42Lefttemporalsuperiorgyrus,BA222946355223.
880.
42Lefttemporalsuperiorgyrus,BA226355183.
680.
39Rightfrontalrectalgyrus,BA11108416263.
880.
42Rightfrontalmiddlegyrus,BA101144159103.
830.
41Righttemporalmiddlegyrus,BA37975458103.
780.
40Leftparietalsuperiorlobule,BA7301867583.
780.
40Righttemporalmiddlegyrus,BA21837332143.
740.
40Leftfrontalmiddlegyrus,BA8233438413.
680.
39Righttemporalmiddlegyrus,BA2177624973.
680.
39Rightfrontalmiddlegyrus,BA634488473.
630.
39Rightfrontalsuperiorgyrus,BA8192623513.
600.
38WhitemattercorrelationswithFSIQLeftfrontalsuperiorgyrus,BA10188206414.
620.
50Rightoccipitalcuneus,BA18338478194.
260.
46Leftfrontalinferiorgyrus,BA45333312994.
230.
46Leftfrontalinferiorgyrus,BA474331134.
020.
43Rightparietalprecuneus,BA7195660464.
060.
44Lefttemporalmiddlegyrus,BA20211574184.
040.
44Righttemporalinferiorgyrus,BA21255812183.
810.
41Righttemporalmiddlegyrus,BA2036543993.
710.
40Leftfrontalprecentralgyrus,BA4425519123.
700.
39Righttemporalmiddlegyrus,BA2111593553.
610.
38x,y,andzcoordinatesareinTalairachatlasspace.
*P<0.
05correctedformultiplecomparisons.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433430twosamplesforthedistributionacrossbrainlobesofthemajorclustersofvoxelswithsignificantcorrelationsbetweengraymatterandFSIQalongwiththetotalnumberofvoxelsintheseclusters.
BothsamplesshowthemostclustersandthehighestnumberofvoxelswhereGMandFSIQarecorrelated(P<0.
001)tobeinthefrontallobes,althoughtheolderUCIsampleshowsahigherportionoffrontallobeclustersthantheyoungerUNMsample(71.
7%versus55.
3%).
TheyoungerUNMsamplehasmoreclustersandvoxelswithaGM/FSIQcorrelationintemporalandlimbiclobes,andtheolderUCIsamplehasmoreintheparietallobes.
ThetotalnumberofvoxelsintheUNMsamplewithaGM/FSIQcorrelation(P<0.
001)is41,303andthisrepresents6.
2%ofalltotalgraymattervoxelsintheaverageUNMsubject(660,870totalGMvoxels).
ThetotalGM/FSIQvoxelsintheolderUCIsampleis31,300or5%ofthetotalgraymattervoxelsintheaverageUCIsubject(644,496totalGMvoxels).
DiscussionThesefindingssupporttheviewthatindividualdifferencesingrayandwhitemattervolumes,inarelativelysmallnumberofareasdistributedthroughoutthebrain,accountforconsiderablevarianceinindividualdifferencesingeneralintelligence.
ThelocationsofourstrongestconjunctionGMfindings(P<0.
05,correctedformultiplecomparisons)inthefrontallobes(BA10,46,and9)areconsistentwithearlierfunctionalimagingfindingsandreinforcetheimpor-tanceoffrontalareasforgeneralintelligence(Duncanetal.
,2000;Grayetal.
,2003;Haieretal.
,1988).
WealsohadsimilarlystrongGMfindingswithposteriorareasincludingBA37,19,40,and43,whichareconsistentwithearlierfunctionalimagingfindingsiden-tifyingthatactivityintheseareaswascorrelatedtogeneralintelli-gence(Duncanetal.
,2000;Grayetal.
,2003;Haieretal.
,1988,2003b).
Interestingly,intheyoungerUMNsample,thestrongestGM/FSIQcorrelationwasinlefttemporallobe,BA37(seeHaieretal.
,2003b).
TheseresultsexpanduponrecentresearchrelatingIQtograymatterinanormalpediatricpopulationusingVBM(Wilkeetal.
,2003).
Theseauthorsfoundgraymatterwithintheanteriorcingu-latetobemostassociatedwithperformanceontheWechslerIntelligenceScaleforChildren—III,withtheolderchildren(meanage=15.
4F1.
86years)accountingforthebulkoftheeffect.
Itisofnotethatouryounggroup(meanage=27F5.
9years)displayednumerouscorrelationsatP<0.
001withinmedialfrontalgrayandwhitematterregionsadjacenttotheanteriorcingulatecortex.
Takentogether,therelationshipbetweenfrontalbrainregionsandIQacrossdifferentagesappearstoprogressfromanteriorcingulate(pediatriccohort),tomedialfrontal(youngUNMcohort),tomoredorsolateralfrontalregions(olderUCIcohort).
Wesuspectthattheprogressionobservedacrossthesecohortsislikelymodulatedbyacceleratedbrainvolumelossacrossthelifespanpreferentiallyaffectingfrontalgraymatter(Razetal.
,1997),particularlytheanteriorcingulategyrus(Goodetal.
,2001).
OurinterpretationofthesedataislimitedbecausedifferentMRIscannerswereusedbetweensamples.
Althoughcorrelationdatadonotspeaktowhysomeindividualshavemoregraymatterinsomeareasthanotherindividuals,GMinmanyoftheareasidentifiedhereshowhighheritability(Thompsonetal.
,2001).
OurdatasupporttheviewthatmostoftheheritableportionofgvariancecanbeaccountedforbyGMinfrontalareas(Thompsonetal.
,2001),andourdataalsoshowthatadditionalportionsofsignificantgvariancecanbeaccountedforbyGMinposteriorareas.
Wholebrainwhitemattermaybemorecorrelatedtointelli-gencethanwholebraingraymatter(Gignacetal.
,2003),butthereislittledataonregionalwhitemattercorrelatesofintelligence.
OurmainWMfindingsaremostlyadjacenttoourGMfindingsandmayrepresentrelativelyfineinterlacingofGMandWMnoteasilydifferentiatedwiththeVBMtechnique.
Alternatively,thewhitematterareas,whichshowcorrelationswithgeneralintelligence,mayalsorepresentpathwaysindependentofGMthatunderliegeneralintelligence.
Forexample,MRIspectroscopyindicatesthataneurometabolite(N-acetylaspartate,NAA)inWMintheleftoccipitoparietalareaiscorrelatedtogeneralintelligenceinnormalsubjects(Jungetal.
,1999a,b)andmayindicatemoremyelinandthefacilitationofneuraltransmissioninthisarea.
Possiblycon-sistentwiththis,ourstrongestWMfinding(P<0.
05,corrected)wasintherightparietalareaclosesttoBA39,contiguouswiththespectroscopicvoxelofinterest.
Thus,brainvolume–IQcorrela-tionsarenotlikelytobeexhaustiveandmaybeconstrainedbymetabolicintegrityattheleveloftheneuron–axon.
TheseobservationsleadustohypothesizethatmorewhitematterinthisareanearBA39mayfacilitatethetransmissionofsensoryinformationfromnumerousposteriorareastothefrontallobes,wheremoregraymatterresultsinbetterprocessingthatmanifestsashigherintelligencescores.
LargersamplemultivariatestudiescanclarifywhichareasworktogetherandaccountformostgvarianceandstudiesofgroupswithimpairedregionalGMmayhelpdeterminewhetherfrontalGMisnecessaryorsufficientforobtaininghighgscores.
AsindicatedinTable1andFig.
2,youngeradultsmayshowdifferentpatternsthanolderadults,likelyreflectingthecumulativeeffectsofneuronallossovertime.
Thefindingthatgrayandwhitemattervolumesinanumberofareassimilarlyaccountforconsiderablevarianceingeneralintel-ligencesuggestsabasisforwhypeopleofthesameIQoftenshowdifferentcognitivestrengthsandweaknesses.
Structuralbraindifferencesingrayandwhitemattervolumesinspecificareasmay,tosomeextent,determinethepatternoffunctionalcorrela-Table4NumberandsizeofvoxelclusterswithasignificantcorrelationbetweengraymatterandFSIQforeachbrainlobeintheUNMandUCIsamplesaFrontalParietalTemporalLimbicOccipitalCerebellumTotalUNM,No.
ofclusters151551027UNM,totalvoxels22,84625114,9013,022283041,303Percentageoftotal55.
3%.
6%36%7.
3%.
7%0UCI,No.
ofclusters136600126UCI,totalvoxels22,4575,5193,0190030531,300Percentageoftotal71.
7%17.
6%9.
6%001%aThesedataaresummarizedfromTables2and3;subclustersarenotincludedinthissummary.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433431tionsinimagingstudiesofintelligenceindependentlyoftaskdemands.
Foranyindividual,thepatternofGMandWMvolumesinrelevantareasmayconstrainwhichareasworktogetherandareactivatedordeactivatedduringproblemsolving,reasoning,orevenpassiveinformationprocessing.
Similartaskperformancemaybeattainedifdifferentcombinationsofbrainareasprovideindepen-dentpathwaysforgoodperformance.
ThiswouldalsosuggestabasisfortheobservationthatregionalbraindamagefromheadinjuryorstrokeoftendoesnotdecreaseIQ.
ThesedataalsosuggestthatitmaybepossibletoassessindividualdifferencesinmentalabilityusingmultivariatecombinationsofgrayandwhitemattervolumesfromarelativelysmallnumberofbrainregionseasilyassessedusingstructuralMRIs.
Havingmoregraymatterinanareaavailableforprocessingmayalsoaccountfortheinversecorrelationsreportedinseveralstudiesbetweenbrainactivationandgoodperformanceong-loadedtasks(Haieretal.
,1988;Parksetal.
,1988),providedthatmoregraymatterresultsinlessenergyusewhenthatareaisemployed(efficiently)forspecificcognitivetasks.
Althoughin-versecorrelationshavebeenreportedbetweenbrainsizeandcerebralglucosemetabolicrate(Haieretal.
,1995;Hatazawaetal.
,1987;Yoshiietal.
,1988),additionalstudyisneededofregionalcorrelationsbetweencerebralstructureandfunction.
Finally,wenotetherelativelysmallproportionofdistributedgraymattervoxelscorrelatedtoFSIQinbothsamples.
Thissuggeststhattheintegrationoftheconstantflowofinformationmovingthroughoutthebrainthatisimportanttointelligentbehaviormaywellbeaccomplishedwithanefficientuseofrelativelyfewandfinitestructures.
Weareremindedbyoneofthefoundingfathersofneuroscience,A.
R.
Luria,thatcomplexbehaviorsare''organizedinsystemsofconcertedlyworkingzones,eachofwhichperformsitsroleincomplexfunctionalsystem,andwhichmaybelocatedincompletelydifferentandoftenfardistantareasofthebrain''(Luria,1973,p.
31).
Ourresearchhighlightsthe''dynamiclocalization''ofintellectualprocessesacrossthelifespanandsuggestsacriticalinterplayofdiscretefrontalandposteriorbrainregions.
AcknowledgmentTheUCIportionofthisworkwasfundedbyagrantfromNICHDtoDr.
Haier(HD037427).
ReferencesAshburner,J.
,Friston,K.
J.
2000.
Voxel-basedmorphometry—Theme-thods.
NeuroImage11(6Pt1),805–821.
Ashburner,J.
,Friston,K.
J.
2001.
Whyvoxel-basedmorphometryshouldbeused.
NeuroImage14(6),1238–1243.
Boivin,M.
J.
,Giordani,B.
,Berent,S.
,etal.
,1992.
Verbalfluencyandpositronemissiontomographicmappingofregionalcerebralglucose-metabolism.
Cortex28(2),231–239.
Duncan,J.
,Seitz,R.
J.
,Kolodny,J.
,etal.
,2000.
Aneuralbasisforgeneralintelligence.
Science289(5478),457–460.
Flashman,L.
A.
,Andreasen,N.
C.
,Flaum,M.
,etal.
,1997.
Intelligenceandregionalbrainvolumesinnormalcontrols.
Intelligence25(3),149–160.
Gignac,G.
,Vernon,P.
A.
,Wickett,J.
C.
,2003.
Factorsinfluencingtherelationshipbetweenbrainsizeandintelligence.
TheScientificStudyofgeneralIntelligence.
H.
Nyborg,Pergamon,Amsterdam,pp.
93–106.
Good,C.
D.
,Johnsrude,I.
S.
,Ashburner,J.
,etal.
,2001.
Avoxel-basedmorphometricstudyofageingin465normaladulthumanbrains.
Neu-roImage14(1Pt1),21–36.
Good,C.
D.
,Scahill,R.
I.
,Fox,N.
C.
,etal.
,2002.
Automaticdifferentiationofanatomicalpatternsinthehumanbrain:validationwithstudiesofdegenerativedementias.
NeuroImage17(1),29–46.
Gray,J.
R.
,Chabris,C.
F.
,Braver,T.
S.
,2003.
Neuralmechanismsofgeneralfluidintelligence.
Nat.
Neurosci.
6(3),316–322.
Haier,R.
J.
,Benbow,C.
P.
,1995.
Sexdifferencesandlateralizationintem-porallobeglucosemetabolismduringmathematicalreasoning.
Dev.
Neuropsychol.
11(4),405–414.
Haier,R.
J.
,Siegel,B.
V.
,Nuechterlein,K.
H.
,etal.
,1988.
Corticalglucosemetabolicratecorrelatesofabstractreasoningandattentionstudiedwithpositronemissiontomography.
Intelligence12(2),199–217.
Haier,R.
J.
,Siegel,B.
V.
Jr.
,MacLachlan,A.
,etal.
,1992a.
Regionalglu-cosemetabolicchangesafterlearningacomplexvisuospatial/motortask:apositronemissiontomographicstudy.
BrainRes.
570(1–2),134–143.
Haier,R.
J.
,Siegel,B.
V.
,Tang,C.
,etal.
,1992b.
Intelligenceandchangesinregionalcerebralglucosemetabolicratefollowinglearning.
Intelligence16(3–4),415–426.
Haier,R.
J.
,Chueh,D.
,Touchette,P.
,etal.
,1995.
BrainsizeandcerebralglucosemetabolicrateinnonspecificmentalretardationandDownsyndrome.
Intelligence20(2),191–210.
Haier,R.
J.
,Alkire,M.
T.
,White,N.
S.
,etal.
,2003a.
Temporalcortexhy-permetabolisminDownsyndromepriortotheonsetofdementia.
Neu-rology61(12),1673–1679.
Haier,R.
J.
,White,N.
S.
,Alkire,M.
T.
,2003b.
Individualdifferencesingeneralintelligencecorrelatewithbrainfunctionduringnonreasoningtasks.
Intelligence31(5),429–441.
Hatazawa,J.
,Brooks,R.
A.
,DiChiro,G.
,etal.
,1987.
Glucoseutilizationrateversusbrainsizeinhumans.
Neurology37(4),583–588.
Jensen,A.
1980.
BiasinMentalTestingFreePress,NewYork.
Jung,R.
E.
,Brooks,W.
M.
,Yeo,R.
A.
,etal.
,1999a.
Biochemicalmarkersofintelligence:aprotonMRspectroscopystudyofnormalhumanbrain.
ProceedingsoftheRoyalSocietyofLondonSeriesB-BiologicalSci-ences266(1426),1375–1379.
Jung,R.
E.
,Yeo,R.
A.
,Chiulli,S.
J.
,etal.
,1999b.
Biochemicalmarkersofcognition:aprotonMRspectroscopystudyofnormalhumanbrain.
NeuroReport10(16),3327–3331.
Luria,A.
R.
,1973.
TheWorkingBrain:AnIntroductiontoNeuropsy-chology.
PenguinPress,London.
MacLullich,A.
M.
J.
,Ferguson,K.
J.
,Deary,I.
J.
,etal.
,2002.
Intracranialcapacityandbrainvolumesareassociatedwithcognitioninhealthyelderlymen.
Neurology59(2),169–174.
Parks,R.
W.
,Loewenstein,D.
A.
,Dodrill,K.
L.
,etal.
,1988.
Cerebralmet-aboliceffectsofaverbalfluencytest—APetScanStudy.
J.
Clin.
Exp.
Neuropsychol.
10(5),565–575.
Prabhakaran,V.
,Smith,J.
A.
,Desmond,J.
E.
,etal.
,1997.
Neuralsubstratesoffluidreasoning:anfMRIstudyofneocorticalactivationduringper-formanceoftheRaven'sProgressiveMatricesTest.
Cogn.
Psychol.
33(1),43–63.
Price,C.
J.
,Friston,K.
J.
,1997.
Cognitiveconjunction:anewapproachtobrainactivationexperiments.
NeuroImage5(4Pt.
1),261–270.
Raz,N.
,Gunning,F.
M.
,Head,D.
,etal.
,1997.
Selectiveagingofthehumancerebralcortexobservedinvivo:differentialvulnerabilityoftheprefrontalgraymatter.
Cereb.
Cortex7(3),268–282.
Spearman,C.
,1904.
Generalintelligenceobjectivelydeterminedandmea-sured.
Am.
J.
Psychol.
15,201–293.
TalairachJ.
,TournouxP.
,1988.
Co-planarstereotaxicatlasofthehumanbrain:a3-dimensionalproportionalsystem,anapproachtocerebralimaging,G.
Thieme,Stuttgart;ThiemeMedicalPublishers,NewYork.
Thompson,P.
M.
,Cannon,T.
D.
,Narr,K.
L.
,etal.
,2001.
Geneticinfluencesonbrainstructure.
Nat.
Neurosci.
4(12),1253–1258.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433432Wechsler,D.
,1981.
WechslerAdultIntelligenceScale—Revised.
Psycho-logicaCorporation,SanAntonio,TX.
White,N.
B.
,Alkire,M.
T.
,Haier,R.
J.
,2003.
Avoxel-basedmorphometricstudyofnon-dementedadultswithDownsyndrome.
NeuroImage20(1),393–403.
Wilke,M.
,Sohn,J.
H.
,Byars,A.
W.
,etal.
,2003.
Brightspots:correlationsofgraymattervolumewithIQinanormalpediatricpopulation.
Neuro-Image20(1),202–215.
Yoshii,F.
,Barker,W.
W.
,Chang,J.
Y.
,etal.
,1988.
Sensitivityofcere-bralglucosemetabolismtoage,gender,brainvolume,brainatrophy,andcerebrovascularriskfactors.
J.
Cereb.
BloodFlowMetab.
8(5),654–661.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433433
Haier,a,*RexE.
Jung,bRonaldA.
Yeo,cKevinHead,aandMichaelT.
AlkiredaDepartmentofPediatrics,UniversityofCalifornia,Irvine,CA92697-5000,USAbDepartmentofNeurology,andtheMINDInstitute,UniversityofNewMexico,Albuquerque,NM87131,USAcDepartmentofPsychology,UniversityofNewMexico,Albuquerque,NM87131,USAdDepartmentofAnesthesiology,UniversityofCaliforniaIrvineMedicalCenter,Orange,CA92868-1350,USAReceived10March2004;revised14April2004;accepted22April2004Availableonline15July2004Totalbrainvolumeaccountsforabout16%ofthevarianceingeneralintelligencescores(IQ),buthowvolumesofspecificregions-of-interest(ROIs)relatetoIQisnotknown.
Weusedvoxel-basedmorphometry(VBM)intwoindependentsamplestoidentifysubstantialgraymatter(GM)correlatesofIQ.
Basedonstatisticalconjunctionofbothsamples(N=47;P<0.
05correctedformultiplecomparisons),moregraymatterisassociatedwithhigherIQindiscreteBrodmannareas(BA)includingfrontal(BA10,46,9),temporal(BA21,37,22,42),parietal(BA43and3),andoccipital(BA19)lobesandnearBA39forwhitematter(WM).
Theseresultsunderscorethedistributedneuralbasisofintelligenceandsuggestadevelopmentalcourseforvolume–IQrelationshipsinadulthood.
D2004ElsevierInc.
Allrightsreserved.
Keywords:IQ;Brainvolume;MorphometryIntroductionCorrelationsbetweenregionalbrainfunctionandperformanceonmentaltestsassociatedwithageneralintelligencefactor(g)asdefinedoriginallybySpearman(1904)havebeendemonstratedmanytimesinnormalsubjects(Duncanetal.
,2000;Grayetal.
,2003;Haieretal.
,1988;HaierandBenbow,1995;Parksetal.
,1988;Prabhakaranetal.
,1997).
Mostofthesestudiesshowthatgoodtestperformancerecruitsareasdistributedthroughoutthebrain,althoughacasehasbeenmadethatactivationwithinareasofthefrontallobesistheprimarysourceofdifferencesing-loadedtestperformance(Duncanetal.
,2000).
Thereisevidencethatdeactivationwithinsomebrainareas,includingfrontallobes,isassociatedwithbettermentaltaskperformance(Haieretal.
,1988,1992a;Parksetal.
,1988),especiallyinsubjectswithhigherintelligencetestscores(Haieretal.
,1992b).
Evenwhenapassivetaskwithnoinherentproblemsolvingisused,subjectswithhigherintelligencescoresshowmoreactivationinposteriorinformationprocessingareasthansubjectswithlowerscores(Boivinetal.
,1992;Haieretal.
,2003b).
Functionalbrainimagingstudiesalwaysmustbeinterpretedtotakeaccountofthespecifictaskdemandsofthementaltaskusedduringtheimagingprotocol.
Thismakesinconsistenciesamongstudyresultsdifficulttoreconcilegiventhewidevarietyoftasksused.
Totheextentthatindividualdifferencesingeneralintelli-gencehaveastructuralcomponent,examiningstructuralcorrelatesofintelligencewouldeliminateanytask-relatedinfluencesfromconsideration.
Forthisreason,structuralimagingofregionalgrayandwhitemattervolumeswouldprovideuniqueinformationaboutthedistributionofbrainareasrelatedtogeneralintelligence.
Forexample,totalbrainvolumeassessedbyMRIinmanystudieshasbeenshowntocorrelateaboutr=0.
40,withintelli-gencescoresandtotalgrayandwhitemattervolumesalsoshowsmallcorrelationswithIQ(Gignacetal.
,2003),butattemptstorelatevolumeofspecificbrainareastotestscoreshavebeenmostlyunsuccessful(Flashmanetal.
,1997;MacLullichetal.
,2002).
Untilnow,suchattemptshaveusedvariousregion-of-interest(ROI)methodsthataredifficulttoreliablyapplytomanybraingyriwhenoutlinedbyhandandoftendonotconformwelltotheextensiveindividualdifferencesamongsubjectsinbrainsizeandmorphologywhenappliedstereotactically.
Arecentmethod-ologicaladvanceisoptimizedvoxel-basedmorphometry(VBM),whichusesalgorithmstosegmentgraymatter(GM)andwhitematter(WM)fromstructuralMRIs(AshburnerandFriston,2000;Goodetal.
,2001).
VBMhasbeenvalidatedextensively(Ash-burnerandFriston,2001;Goodetal.
,2002)andithasbeenused,forexample,tocharacterizegrayandwhitemattervolumechangesinaging(Goodetal.
,2001),dementia(Goodetal.
,2002),andDownsyndrome(Whiteetal.
,2003).
MaterialsandmethodsSubjectsWetestedtwosamplesandusedastatisticalconjunctionapproach(PriceandFriston,1997)toshowwherecorrelationsbetweenIQandgrayorwhitematterwerecommontobothsamples.
Thefirstsamplewas23normalvolunteers(14womenand9men;1053-8119/$-seefrontmatterD2004ElsevierInc.
Allrightsreserved.
doi:10.
1016/j.
neuroimage.
2004.
04.
025*Correspondingauthor.
DepartmentofPediatrics,UniversityofCaliforniaIrvine,MedicalScienceI,B140,Irvine,CA92697.
Fax:+1-949-824-9059.
E-mailaddress:rjhaier@uci.
edu(R.
J.
Haier).
AvailableonlineonScienceDirect(www.
sciencedirect.
com.
)www.
elsevier.
com/locate/ynimgNeuroImage23(2004)425–433meanage=27,SD=5.
9,range=18–37)recruitedfromtheUniversityofNewMexico(UNM).
Sample1MRIswereobtainedwitha1.
5-Tscanner,headcoil,andsoftware(Signa5.
4;GeneralElectricMedicalSystems,Waukesha,WI).
AT1sagittallocalizersequence(TE=6.
9ms,TR=200ms,FOV=2424cm2,fiveslices,thickness=5mm,spacing=2.
5mm,matrix=256128)wasacquired,followedbyaT1-weightedaxialseries(fastRFspoiledgradient-recalled,TE=6.
9ms,TR=17.
7ms,flipangle=25j,matrix=256192,120slices,thickness=1.
5mm)togivefullbraincoverage.
Thesecondsamplewas24normalvolunteers(13menand11women,meanage=59,SD=15.
9,range37–84)recruitedattheUniversityofCalifornia(UCI),Irvine,asmiddle-agedandoldernormalcontrolsforanimagingstudyofdementiainDownsyndromeandAlzheimer'sdisease(Haieretal.
,2003a).
Sample2MRIswereobtainedwitha1.
5-TclinicalPhillipsEclipsescanner(PhilipsMedicalSystems,N.
A.
,Bothell,WA).
WeusedT1-weighted,volumetricSPGRMRIscans(FOV=24cm,flipangle=40,TR=24,TE=5).
Theimagesconsistedof120contiguous1.
2-mmthickaxialslices,eachwithanin-planeimagematrixof256256imageelements.
Allimagesinbothsampleswerevisuallyinspectedtoensureimagequality.
IntelligencetestingToassessgeneralintelligence,subjectsweretestedwiththeWechslerAdultIntelligenceScale(WAIS).
TheWAISbattery(Wechsler,1981)consistsof11diversesubtests,whichtapavarietyofverbalandnonverbalmentalabilitiesthatcontributetogeneralintelligence.
TheWAISFullScaleIQ(FSIQ)scoreisbasedonperformanceofall11subtests(accordingtoagebasednorms).
Factoranalyticstudies(Jensen,1980)showthateachsubtestloadsonthegfactorandtheFSIQscoreloadsthehighest(about0.
90or81%ofthevarianceing).
Forthisreason,FSIQisconsideredoneofthebestindexesofindividualdifferencesingeneralintelligence.
Forsample1(UMN),theFSIQwas116(SD=14.
7)andtherangewas90–155.
Forsample2(UCI),themeanFSIQwas116(SD=14.
2)andtherangewas90–142.
Voxel-basedmorphometryWeappliedVBMtoidentifybrainareaswhereGMandWMvolumesarecorrelatedtoFSIQtotestwhetheranysuchareasareclusteredinfrontallobesordistributedthroughoutthebrain.
WeusedStatisticalParametricMappingsoftware(SPM2;TheWellcomeDepartmentofImagingNeuroscience,UniversityCollegeLondon)tocreateastudy-specifictemplateandthenappliedtheoptimizedVBMprotocoltoeachsampleseparatelyusingthemethodsofAshburnerandFriston(2000)andGoodetal.
(2001).
Topreservetheamountoftissueinanygivenanatomicalregionafterspatialnormalization,theoptimalGMandWMpartitionsweremultipliedbytheJacobiandetermi-nantsoftheirrespectivespatialtransformationmatrix.
ThereasonforperformingthismodulationstepissothatthefinalVBMstatisticswillreflectlocaldeviationsintheabsoluteamount(volume)oftissueindifferentregionsofthebrain(AshburnerandFriston,2000).
ThemodulatedGMandWMpartitionswerethensmoothedwitha12-mmFWHMisotropicGaussiankerneltoaccountforslightmisalignmentsofhomol-ogousanatomicalstructuresandtoensurestatisticalvalidityunderparametricassumptions.
StatisticalconjunctionapproachWespecificallytestedwhetherregionalgrayandwhitemattervolumeswerecorrelatedwithFSIQscorestreatinganyeffectsofage,sex,andhandedness(threecasesintheUNMsamplewerelefthanded)asnuisancevariablesintheSPM2designmatrix.
Aftercomputingthecorrelationanalysesforeachsampleseparately,weFig.
1.
CorrelationsbetweengrayandwhitematterandFSIQconjunctedacrossbothsamples.
Basedonstatisticalconjunction,correlationsbetweengraymatterandFSIQareshownintheleftcolumnongraymattertemplates;whitemattercorrelationstoFSIQareshownintherightcolumnonawhitemattertemplate.
Toprowshowsfrontalview,secondrowshowsmedialsagittalview,nextrowsshowleftandrightlateralviews,andbottomrowshowsoccipitalview.
Anatomicallocations,atlascoordinates,andclustersizesarelistedinTable1.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433426Table1LocalizationofconjunctioncorrelationsfromUNMandUCIbetweengrayandwhitematterwithFSIQ(PV0.
001)Brainarea(and/ornearestgraymatter)Clustersizexyztr2GraymattercorrelationswithFSIQRightmedialfrontalgyrus,BA10*684626664.
650.
35Rightmedialfrontalgyrus,BA10*863174.
380.
32Rightsuperiorfrontalgyrus,BA9*456314.
290.
31Leftmiddlefrontalgyrus,BA46*34645039174.
320.
31Leftinferiorfrontalgyrus,BA45*513154.
130.
29Leftmiddlefrontalgyrus,BA46*5035243.
920.
27Leftmiddletemporalgyrus,BA21*692664543.
930.
27Rightfrontalprecentralgyrus,BA9*7053723343.
860.
27Rightmiddlefrontalgyrus,BA9*4932323.
220.
20Righttemporalinferiorgyrus,BA37*22945560103.
840.
26Righttemporalmiddlegyrus,BA37*575183.
650.
25Righttemporalmiddlegyrus,BA21*614833.
510.
23Leftmiddlefrontalgyrus,BA10*524405633.
750.
26Leftsuperiorfrontalgyrus,BA10*3651233.
650.
25Leftmiddlefrontalgyrus,BA10*415393.
510.
23Leftmiddleoccipitalgyrus,BA19*345546853.
660.
25Lefttemporalsuperiorgyrus,BA22*4266348193.
610.
24Leftparietalsupramarginalgyrus,BA40*6645263.
420.
22Lefttemporaltransversegyrus,BA42*218629123.
470.
23Rightparietalpostcentralgyrus,BA43*3856417153.
460.
23Rightparietalpostcentralgyrus,BA3*6115233.
360.
22Righttemporalmiddlegyrus,BA21*456535103.
300.
21Leftfrontalsuperiorgyrus,BA8*134123503.
280.
21Rightfrontalinferiorgyrus,BA9*20587303.
280.
21WhitemattercorrelationswithFSIQRighttemporalmiddlegyrus,BA39*4457515453.
480.
23Righttemporalfusiformgyrus,BA375037103.
240.
20Rightmiddletemporalgyrus,BA21553332.
510.
13Lefttemporalmiddlegyrus,BA204325524093.
380.
22Lefttemporalinferiorgyrus,BA19445313.
140.
19Lefttemporalmiddlegyrus,BA37525322.
700.
15Leftfrontalsuperiorgyrus,BA101839186014.
550.
34Leftfrontalmedialgyrus,BA1076383.
920.
27Leftfrontalmedialgyrus,BA10862213.
470.
23Leftfrontalinferiorgyrus,BA478132313132.
980.
18Rightfrontalmiddlegyrus,BA102126295982.
930.
17Rightfrontalsuperiorgyrus,BA10216262.
850.
17Rightfrontalmedialgyrus,BA102444102.
360.
12Leftparietalprecuneus,BA726752360302.
790.
16Leftparietalprecuneus,BA71552242.
370.
12Rightparietalprecuneus,BA3112881858352.
790.
16Rightlimbiccingulategyrus,BA2418191315322.
750.
16Rightlimbiccingulategyrus,BA32139362.
300.
11Rightfrontalmedialgyrus,BA81131392.
240.
11Leftfrontalsuperiorgyrus,BA95821052352.
670.
15Leftfrontalsuperiorgyrus,BA9958322.
170.
10Rightlimbic,parahippocampus,BA357261726212.
550.
14Rightlimbic,parahippocampus,BA351930142.
360.
12Leftlimbic,parahippocampus,BA3515801428192.
460.
13Leftbrainstem,pons431222.
270.
11Leftbrainstem,midbrainsubstanianigra1127112.
210.
11Rightfrontalsuperiorgyrus,BA113242949152.
410.
12Rightfrontalsuperiorgyrus,BA112456152.
110.
10Leftsublobar,insula,BA13245438122.
400.
12Rightcerebellum,anteriorlobe4063046302.
360.
12Rightfrontalsuperiorgyrus,BA63135542.
200.
11Leftfrontalsuperiorgyrus,BA111402047172.
200.
11Leftfrontalprecentralgyrus,BA660491112.
180.
10Righttemporal,subgyral,BA20894216172.
170.
10Leftfrontalsuperiorgyrus,BA658920532.
140.
10x,y,andzcoordinatesareinTalairachatlasspace.
*P<0.
05correctedformultiplecomparisons.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433427usedtheconjunctionapproach(PriceandFriston,1997)toshowwheregraymatter(andwhitematter)correlationsoverlappedfortheUNMandtheUCIsamples(i.
e.
,voxelswithcorrelationsincommonforbothsamples).
Thisconjunctionapproachminimizespotentialproblemsassociatedwithcombiningdatafromdifferentscanners.
Theconjunctionanalysisalsohastheadvantageofmaximizingstatisticalpowerbecauseall47subjectsareusedintheanalysisanditisequivalenttoafixedfactormodelinSPM.
FindingsareconsideredsignificantatP<0.
05correctedformultiplecomparisons;findingsatPV0.
001uncorrectedalsoareshownforhypothesisgeneration.
R2estimatesweredeterminedusingtheformula:R2=t2/(df+t2)wheredf=41(fromtheSPM2conjunctiondesignmatrix).
Locationsofsignificantclusters(cent-roids)areconvertedfromMontrealNeurologicalInstitute(MNI)toTalairachatlas(TalairachandTournoux,1988)coordinatesandreportedasclosestBrodmannarea(BA)wherepossible.
Onlyclustersofatleast10voxelsarereported.
ResultsTheconjunctionresults(N=47;Fig.
1)showedrobustpositivecorrelations(P<0.
05,correctedformultiplecomparisons)betweenFSIQandgraymattervolumesinBrodmannareas(BA)10,46,and9infrontallobes;BA21,37,22,and42intemporallobes;BA43and3inparietallobes;andBA19intheoccipitallobe.
ThesizeandFig.
2.
CorrelationsbetweengrayandwhitematterandFSIQforeachsample.
Basedonseparateanalysesforeachsample,graymattercorrelationsareshownintheleftcolumns(UNMandUCI,respectively);whitemattercorrelationsareshownintherightcolumns.
Toprowshowsfrontalview,secondrowshowsmedialsagittalview,nextrowsshowleftandrightlateralviews,andbottomrowshowsoccipitalview.
Anatomicallocations,atlascoordinates,andclustersizesarelistedinTables2(UNMdata)and3(UCIdata).
R.
J.
Haieretal.
/NeuroImage23(2004)425–433428locationsoftheseareasareshowninTable1.
Similarbutlessrobustcorrelationswithwhitematterareas(PV0.
001,uncorrected)werealsofound(Table1).
Notably,24graymatterregionsweresigni-ficantatP<0.
05(corrected),yetonlyonewhitematterregionwassignificantatthislevel.
EstimatesofFSIQvariabilityaccountedforbythehighestcorrelationforanindividualvoxelwithinthesignificantgrayandwhitematterareas(R2)areashighas73%(Table1).
Therewerenosignificantnegativecorrelations.
Table2LocalizationofcorrelationsintheUNMsamplebetweengrayandwhitematterwithFSIQ(PV0.
001)Brainarea(and/ornearestgraymatter)Clustersizexyztr2GraymattercorrelationswithFSIQLefttemporalfusiformgyrus,BA37*3984415296.
930.
73Lefttemporalfusiformgyrus,BA364439274.
170.
49Lefttemporalfusiformgyrus,BA363943234.
090.
48Leftfrontalsuperiorgyrus,BA1018,470760256.
280.
69Rightfrontalsuperiorgyrus,BA10763186.
030.
67Rightfrontalsuperiorgyrus,BA8943545.
510.
63Leftfrontalsuperiorgyrus,BA61086017625.
990.
67Righttemporalfusiformgyrus,BA3610,1874436305.
940.
66Righttemporalfusiformgyrus,BA205140275.
290.
61Righttemporalmiddlegyrus,BA37576215.
180.
60Rightlimbiclobe,anteriorcingulate,BA321756143075.
360.
61Rightlimbiclobe,anteriorcingulate,BA321731174.
420.
52Rightfrontalinferiorgyrus,BA479623113205.
230.
60Leftparietalangulargyrus,BA392513058344.
810.
56Leftlimbiclobe,anteriorcingulate,BA328301738164.
780.
56Leftfrontalmiddlegyrus,BA10602405874.
710.
55Leftfrontalmiddlegyrus,BA103952133.
930.
46Rightfrontalsuperiorgyrus,BA111213046224.
660.
55Leftsublobar,insula,BA13151743344.
630.
54Leftsublobar,insula,BA13371374.
000.
47Leftoccipitalmiddlegyrus,BA19283616594.
480.
53Leftlimbiclobe,cingulategyrus,BA23338816304.
210.
50Leftlimbiclobe,cingulategyrus,BA2496323.
730.
44Rightmedialfrontalgyrus,BA10275135144.
160.
49Leftmiddletemporalgyrus,BA22597514714.
160.
49Leftfrontalinferiorgyrus,BA476252924194.
150.
49Leftfrontalinferiorgyrus,BA473828164.
100.
48Leftfrontalinferiorgyrus,BA472516183.
910.
46Lefttemporalfusiformgyrus,BA201136011284.
030.
47Rightfrontalinferiorgyrus,BA92725421243.
970.
47Leftfrontalmiddlegyrus,BA461854841203.
910.
46Leftfrontalmiddlegyrus,BA8174224493.
890.
46Rightfrontalmiddlegyrus,BA8493830493.
890.
46Leftlimbiclobe,anteriorcingulate,BA10691232113.
850.
45Rightfrontalmiddlegyrus,BA10334240183.
830.
45Rightfrontalmiddlegyrus,BA9944121343.
790.
44Rightlimbiclobe,parahippocampus,BA1929344503.
790.
44Leftfrontalmedialgyrus,BA32201414443.
750.
44Leftinferiorfrontalgyrus,BA4735542133.
740.
44Lefttemporalsuperiorgyrus,BA38203613203.
710.
43WhitemattercorrelationswithFSIQLeftfrontalsuperiorgyrus,BA9690850215.
870.
66Leftfrontalmedialgyrus,BA9844293.
840.
45Rightfrontalmedialgyrus,BA102234845145.
190.
60Rightfrontalmedialgyrus,BA101847114.
640.
54Rightfrontalsuperior,BA102555134.
070.
48Rightlimbiclobe,parahippocampus,amygdala245287225.
080.
59Rightfrontalsuperiorgyrus,BA84801128485.
010.
58Rightmiddletemporalgyrus,BA21148595244.
950.
58Rightsublobar,insula,BA221138442714.
530.
53Lefttemporalmiddlegyrus,BA202055037104.
320.
51Leftmedialfrontalgyrus,BA10345105754.
140.
49Leftfrontalsuperiorgyrus,BA1076023.
830.
45Rightbrainstem,pons1461421263.
970.
47x,y,andzcoordinatesareinTalairachatlasspace.
*P<0.
05correctedformultiplecomparisons.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433429ThepositivecorrelationsbetweengrayandwhitematterandFSIQforeachsampleseparatelyareshowninFig.
2andinTables2(UNM)and3(UCI);therewerenosignificantnegativecorre-lations.
StatisticalcomparisonsmadedirectlybetweenthetwosamplescouldbedifficulttointerpretbecausedifferentMRIscannerswereusedandtheagesweredifferent.
Anexploratoryanalysisshowedfewareaswithsignificantdifferencesbetweengrayandwhitemattervolumesbetweenthesamples,andnoneoftheseareasoverlappedwithareasidentifiedintheconjunctionanalysis.
AnotherapproachtoassessingthedifferencesbetweenthesamplesisshowninTable4.
ItshowsacomparisonbetweentheTable3LocalizationofcorrelationsintheUCIsamplebetweengrayandwhitematterwithFSIQ(PV0.
001)Brainarea(and/ornearestgraymatter)Clustersizexyztr2GraymattercorrelationswithFSIQLeftfrontalinferiorgyrus,BA45*11,086553017.
060.
70Leftfrontalinferiorgyrus,BA465632105.
500.
59Leftfrontalmiddlegyrus,BA10425535.
350.
58Lefttemporalmiddlegyrus,BA212424644535.
150.
56Leftoccipitalmiddlegyrus,BA19536734.
370.
48Lefttemporalmiddlegyrus,BA21673393.
940.
43Rightfrontalprecentralgyrus,BA951773723345.
130.
56Rightfrontalmiddlegyrus,BA465332214.
330.
47Rightfrontalmiddlegyrus,BA94525334.
310.
47Rightfrontalmedialgyrus,BA10186926665.
030.
55Leftfrontalmedialgyrus,BA10565114.
430.
48Rightfrontalsuperiorgyrus,BA102266114.
270.
46Rightfrontalprecentralgyrus,BA63096607414.
830.
53Rightfrontalprecentralgyrus,BA66510324.
230.
46Rightparietalpostcentralgyrus,BA436413204.
180.
45Leftlingualgyrus4378117714.
720.
51Leftoccipitalfusiformgyrus,BA182187124.
180.
45Leftoccipitalcuneus,BA1809664.
010.
43Rightparietalprecuneus,BA7433961644.
580.
50Rightfrontalinferiorgyrus,BA46296363464.
400.
48Leftcerebellum,posteriorlobe3053276224.
120.
45Leftparietalinferiorlobule,BA404875559394.
100.
44Leftparietalinferiorlobule,BA405450463.
790.
41Rightparietalinferiorlobule,BA401115143504.
070.
44Leftparietalpostcentralgyrus,BA780953704.
060.
44Rightfrontalsuperiorgyrus,BA9269458284.
000.
43Rightfrontalmedialgyrus,BA10462203.
730.
40Leftfrontalmiddlegyrus,BA93045216253.
990.
43Lefttemporalmiddlegyrus,BA21446910173.
920.
42Leftfrontalsuperiorgyrus,BA10623651233.
910.
42Lefttemporalsuperiorgyrus,BA222946355223.
880.
42Lefttemporalsuperiorgyrus,BA226355183.
680.
39Rightfrontalrectalgyrus,BA11108416263.
880.
42Rightfrontalmiddlegyrus,BA101144159103.
830.
41Righttemporalmiddlegyrus,BA37975458103.
780.
40Leftparietalsuperiorlobule,BA7301867583.
780.
40Righttemporalmiddlegyrus,BA21837332143.
740.
40Leftfrontalmiddlegyrus,BA8233438413.
680.
39Righttemporalmiddlegyrus,BA2177624973.
680.
39Rightfrontalmiddlegyrus,BA634488473.
630.
39Rightfrontalsuperiorgyrus,BA8192623513.
600.
38WhitemattercorrelationswithFSIQLeftfrontalsuperiorgyrus,BA10188206414.
620.
50Rightoccipitalcuneus,BA18338478194.
260.
46Leftfrontalinferiorgyrus,BA45333312994.
230.
46Leftfrontalinferiorgyrus,BA474331134.
020.
43Rightparietalprecuneus,BA7195660464.
060.
44Lefttemporalmiddlegyrus,BA20211574184.
040.
44Righttemporalinferiorgyrus,BA21255812183.
810.
41Righttemporalmiddlegyrus,BA2036543993.
710.
40Leftfrontalprecentralgyrus,BA4425519123.
700.
39Righttemporalmiddlegyrus,BA2111593553.
610.
38x,y,andzcoordinatesareinTalairachatlasspace.
*P<0.
05correctedformultiplecomparisons.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433430twosamplesforthedistributionacrossbrainlobesofthemajorclustersofvoxelswithsignificantcorrelationsbetweengraymatterandFSIQalongwiththetotalnumberofvoxelsintheseclusters.
BothsamplesshowthemostclustersandthehighestnumberofvoxelswhereGMandFSIQarecorrelated(P<0.
001)tobeinthefrontallobes,althoughtheolderUCIsampleshowsahigherportionoffrontallobeclustersthantheyoungerUNMsample(71.
7%versus55.
3%).
TheyoungerUNMsamplehasmoreclustersandvoxelswithaGM/FSIQcorrelationintemporalandlimbiclobes,andtheolderUCIsamplehasmoreintheparietallobes.
ThetotalnumberofvoxelsintheUNMsamplewithaGM/FSIQcorrelation(P<0.
001)is41,303andthisrepresents6.
2%ofalltotalgraymattervoxelsintheaverageUNMsubject(660,870totalGMvoxels).
ThetotalGM/FSIQvoxelsintheolderUCIsampleis31,300or5%ofthetotalgraymattervoxelsintheaverageUCIsubject(644,496totalGMvoxels).
DiscussionThesefindingssupporttheviewthatindividualdifferencesingrayandwhitemattervolumes,inarelativelysmallnumberofareasdistributedthroughoutthebrain,accountforconsiderablevarianceinindividualdifferencesingeneralintelligence.
ThelocationsofourstrongestconjunctionGMfindings(P<0.
05,correctedformultiplecomparisons)inthefrontallobes(BA10,46,and9)areconsistentwithearlierfunctionalimagingfindingsandreinforcetheimpor-tanceoffrontalareasforgeneralintelligence(Duncanetal.
,2000;Grayetal.
,2003;Haieretal.
,1988).
WealsohadsimilarlystrongGMfindingswithposteriorareasincludingBA37,19,40,and43,whichareconsistentwithearlierfunctionalimagingfindingsiden-tifyingthatactivityintheseareaswascorrelatedtogeneralintelli-gence(Duncanetal.
,2000;Grayetal.
,2003;Haieretal.
,1988,2003b).
Interestingly,intheyoungerUMNsample,thestrongestGM/FSIQcorrelationwasinlefttemporallobe,BA37(seeHaieretal.
,2003b).
TheseresultsexpanduponrecentresearchrelatingIQtograymatterinanormalpediatricpopulationusingVBM(Wilkeetal.
,2003).
Theseauthorsfoundgraymatterwithintheanteriorcingu-latetobemostassociatedwithperformanceontheWechslerIntelligenceScaleforChildren—III,withtheolderchildren(meanage=15.
4F1.
86years)accountingforthebulkoftheeffect.
Itisofnotethatouryounggroup(meanage=27F5.
9years)displayednumerouscorrelationsatP<0.
001withinmedialfrontalgrayandwhitematterregionsadjacenttotheanteriorcingulatecortex.
Takentogether,therelationshipbetweenfrontalbrainregionsandIQacrossdifferentagesappearstoprogressfromanteriorcingulate(pediatriccohort),tomedialfrontal(youngUNMcohort),tomoredorsolateralfrontalregions(olderUCIcohort).
Wesuspectthattheprogressionobservedacrossthesecohortsislikelymodulatedbyacceleratedbrainvolumelossacrossthelifespanpreferentiallyaffectingfrontalgraymatter(Razetal.
,1997),particularlytheanteriorcingulategyrus(Goodetal.
,2001).
OurinterpretationofthesedataislimitedbecausedifferentMRIscannerswereusedbetweensamples.
Althoughcorrelationdatadonotspeaktowhysomeindividualshavemoregraymatterinsomeareasthanotherindividuals,GMinmanyoftheareasidentifiedhereshowhighheritability(Thompsonetal.
,2001).
OurdatasupporttheviewthatmostoftheheritableportionofgvariancecanbeaccountedforbyGMinfrontalareas(Thompsonetal.
,2001),andourdataalsoshowthatadditionalportionsofsignificantgvariancecanbeaccountedforbyGMinposteriorareas.
Wholebrainwhitemattermaybemorecorrelatedtointelli-gencethanwholebraingraymatter(Gignacetal.
,2003),butthereislittledataonregionalwhitemattercorrelatesofintelligence.
OurmainWMfindingsaremostlyadjacenttoourGMfindingsandmayrepresentrelativelyfineinterlacingofGMandWMnoteasilydifferentiatedwiththeVBMtechnique.
Alternatively,thewhitematterareas,whichshowcorrelationswithgeneralintelligence,mayalsorepresentpathwaysindependentofGMthatunderliegeneralintelligence.
Forexample,MRIspectroscopyindicatesthataneurometabolite(N-acetylaspartate,NAA)inWMintheleftoccipitoparietalareaiscorrelatedtogeneralintelligenceinnormalsubjects(Jungetal.
,1999a,b)andmayindicatemoremyelinandthefacilitationofneuraltransmissioninthisarea.
Possiblycon-sistentwiththis,ourstrongestWMfinding(P<0.
05,corrected)wasintherightparietalareaclosesttoBA39,contiguouswiththespectroscopicvoxelofinterest.
Thus,brainvolume–IQcorrela-tionsarenotlikelytobeexhaustiveandmaybeconstrainedbymetabolicintegrityattheleveloftheneuron–axon.
TheseobservationsleadustohypothesizethatmorewhitematterinthisareanearBA39mayfacilitatethetransmissionofsensoryinformationfromnumerousposteriorareastothefrontallobes,wheremoregraymatterresultsinbetterprocessingthatmanifestsashigherintelligencescores.
LargersamplemultivariatestudiescanclarifywhichareasworktogetherandaccountformostgvarianceandstudiesofgroupswithimpairedregionalGMmayhelpdeterminewhetherfrontalGMisnecessaryorsufficientforobtaininghighgscores.
AsindicatedinTable1andFig.
2,youngeradultsmayshowdifferentpatternsthanolderadults,likelyreflectingthecumulativeeffectsofneuronallossovertime.
Thefindingthatgrayandwhitemattervolumesinanumberofareassimilarlyaccountforconsiderablevarianceingeneralintel-ligencesuggestsabasisforwhypeopleofthesameIQoftenshowdifferentcognitivestrengthsandweaknesses.
Structuralbraindifferencesingrayandwhitemattervolumesinspecificareasmay,tosomeextent,determinethepatternoffunctionalcorrela-Table4NumberandsizeofvoxelclusterswithasignificantcorrelationbetweengraymatterandFSIQforeachbrainlobeintheUNMandUCIsamplesaFrontalParietalTemporalLimbicOccipitalCerebellumTotalUNM,No.
ofclusters151551027UNM,totalvoxels22,84625114,9013,022283041,303Percentageoftotal55.
3%.
6%36%7.
3%.
7%0UCI,No.
ofclusters136600126UCI,totalvoxels22,4575,5193,0190030531,300Percentageoftotal71.
7%17.
6%9.
6%001%aThesedataaresummarizedfromTables2and3;subclustersarenotincludedinthissummary.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433431tionsinimagingstudiesofintelligenceindependentlyoftaskdemands.
Foranyindividual,thepatternofGMandWMvolumesinrelevantareasmayconstrainwhichareasworktogetherandareactivatedordeactivatedduringproblemsolving,reasoning,orevenpassiveinformationprocessing.
Similartaskperformancemaybeattainedifdifferentcombinationsofbrainareasprovideindepen-dentpathwaysforgoodperformance.
ThiswouldalsosuggestabasisfortheobservationthatregionalbraindamagefromheadinjuryorstrokeoftendoesnotdecreaseIQ.
ThesedataalsosuggestthatitmaybepossibletoassessindividualdifferencesinmentalabilityusingmultivariatecombinationsofgrayandwhitemattervolumesfromarelativelysmallnumberofbrainregionseasilyassessedusingstructuralMRIs.
Havingmoregraymatterinanareaavailableforprocessingmayalsoaccountfortheinversecorrelationsreportedinseveralstudiesbetweenbrainactivationandgoodperformanceong-loadedtasks(Haieretal.
,1988;Parksetal.
,1988),providedthatmoregraymatterresultsinlessenergyusewhenthatareaisemployed(efficiently)forspecificcognitivetasks.
Althoughin-versecorrelationshavebeenreportedbetweenbrainsizeandcerebralglucosemetabolicrate(Haieretal.
,1995;Hatazawaetal.
,1987;Yoshiietal.
,1988),additionalstudyisneededofregionalcorrelationsbetweencerebralstructureandfunction.
Finally,wenotetherelativelysmallproportionofdistributedgraymattervoxelscorrelatedtoFSIQinbothsamples.
Thissuggeststhattheintegrationoftheconstantflowofinformationmovingthroughoutthebrainthatisimportanttointelligentbehaviormaywellbeaccomplishedwithanefficientuseofrelativelyfewandfinitestructures.
Weareremindedbyoneofthefoundingfathersofneuroscience,A.
R.
Luria,thatcomplexbehaviorsare''organizedinsystemsofconcertedlyworkingzones,eachofwhichperformsitsroleincomplexfunctionalsystem,andwhichmaybelocatedincompletelydifferentandoftenfardistantareasofthebrain''(Luria,1973,p.
31).
Ourresearchhighlightsthe''dynamiclocalization''ofintellectualprocessesacrossthelifespanandsuggestsacriticalinterplayofdiscretefrontalandposteriorbrainregions.
AcknowledgmentTheUCIportionofthisworkwasfundedbyagrantfromNICHDtoDr.
Haier(HD037427).
ReferencesAshburner,J.
,Friston,K.
J.
2000.
Voxel-basedmorphometry—Theme-thods.
NeuroImage11(6Pt1),805–821.
Ashburner,J.
,Friston,K.
J.
2001.
Whyvoxel-basedmorphometryshouldbeused.
NeuroImage14(6),1238–1243.
Boivin,M.
J.
,Giordani,B.
,Berent,S.
,etal.
,1992.
Verbalfluencyandpositronemissiontomographicmappingofregionalcerebralglucose-metabolism.
Cortex28(2),231–239.
Duncan,J.
,Seitz,R.
J.
,Kolodny,J.
,etal.
,2000.
Aneuralbasisforgeneralintelligence.
Science289(5478),457–460.
Flashman,L.
A.
,Andreasen,N.
C.
,Flaum,M.
,etal.
,1997.
Intelligenceandregionalbrainvolumesinnormalcontrols.
Intelligence25(3),149–160.
Gignac,G.
,Vernon,P.
A.
,Wickett,J.
C.
,2003.
Factorsinfluencingtherelationshipbetweenbrainsizeandintelligence.
TheScientificStudyofgeneralIntelligence.
H.
Nyborg,Pergamon,Amsterdam,pp.
93–106.
Good,C.
D.
,Johnsrude,I.
S.
,Ashburner,J.
,etal.
,2001.
Avoxel-basedmorphometricstudyofageingin465normaladulthumanbrains.
Neu-roImage14(1Pt1),21–36.
Good,C.
D.
,Scahill,R.
I.
,Fox,N.
C.
,etal.
,2002.
Automaticdifferentiationofanatomicalpatternsinthehumanbrain:validationwithstudiesofdegenerativedementias.
NeuroImage17(1),29–46.
Gray,J.
R.
,Chabris,C.
F.
,Braver,T.
S.
,2003.
Neuralmechanismsofgeneralfluidintelligence.
Nat.
Neurosci.
6(3),316–322.
Haier,R.
J.
,Benbow,C.
P.
,1995.
Sexdifferencesandlateralizationintem-porallobeglucosemetabolismduringmathematicalreasoning.
Dev.
Neuropsychol.
11(4),405–414.
Haier,R.
J.
,Siegel,B.
V.
,Nuechterlein,K.
H.
,etal.
,1988.
Corticalglucosemetabolicratecorrelatesofabstractreasoningandattentionstudiedwithpositronemissiontomography.
Intelligence12(2),199–217.
Haier,R.
J.
,Siegel,B.
V.
Jr.
,MacLachlan,A.
,etal.
,1992a.
Regionalglu-cosemetabolicchangesafterlearningacomplexvisuospatial/motortask:apositronemissiontomographicstudy.
BrainRes.
570(1–2),134–143.
Haier,R.
J.
,Siegel,B.
V.
,Tang,C.
,etal.
,1992b.
Intelligenceandchangesinregionalcerebralglucosemetabolicratefollowinglearning.
Intelligence16(3–4),415–426.
Haier,R.
J.
,Chueh,D.
,Touchette,P.
,etal.
,1995.
BrainsizeandcerebralglucosemetabolicrateinnonspecificmentalretardationandDownsyndrome.
Intelligence20(2),191–210.
Haier,R.
J.
,Alkire,M.
T.
,White,N.
S.
,etal.
,2003a.
Temporalcortexhy-permetabolisminDownsyndromepriortotheonsetofdementia.
Neu-rology61(12),1673–1679.
Haier,R.
J.
,White,N.
S.
,Alkire,M.
T.
,2003b.
Individualdifferencesingeneralintelligencecorrelatewithbrainfunctionduringnonreasoningtasks.
Intelligence31(5),429–441.
Hatazawa,J.
,Brooks,R.
A.
,DiChiro,G.
,etal.
,1987.
Glucoseutilizationrateversusbrainsizeinhumans.
Neurology37(4),583–588.
Jensen,A.
1980.
BiasinMentalTestingFreePress,NewYork.
Jung,R.
E.
,Brooks,W.
M.
,Yeo,R.
A.
,etal.
,1999a.
Biochemicalmarkersofintelligence:aprotonMRspectroscopystudyofnormalhumanbrain.
ProceedingsoftheRoyalSocietyofLondonSeriesB-BiologicalSci-ences266(1426),1375–1379.
Jung,R.
E.
,Yeo,R.
A.
,Chiulli,S.
J.
,etal.
,1999b.
Biochemicalmarkersofcognition:aprotonMRspectroscopystudyofnormalhumanbrain.
NeuroReport10(16),3327–3331.
Luria,A.
R.
,1973.
TheWorkingBrain:AnIntroductiontoNeuropsy-chology.
PenguinPress,London.
MacLullich,A.
M.
J.
,Ferguson,K.
J.
,Deary,I.
J.
,etal.
,2002.
Intracranialcapacityandbrainvolumesareassociatedwithcognitioninhealthyelderlymen.
Neurology59(2),169–174.
Parks,R.
W.
,Loewenstein,D.
A.
,Dodrill,K.
L.
,etal.
,1988.
Cerebralmet-aboliceffectsofaverbalfluencytest—APetScanStudy.
J.
Clin.
Exp.
Neuropsychol.
10(5),565–575.
Prabhakaran,V.
,Smith,J.
A.
,Desmond,J.
E.
,etal.
,1997.
Neuralsubstratesoffluidreasoning:anfMRIstudyofneocorticalactivationduringper-formanceoftheRaven'sProgressiveMatricesTest.
Cogn.
Psychol.
33(1),43–63.
Price,C.
J.
,Friston,K.
J.
,1997.
Cognitiveconjunction:anewapproachtobrainactivationexperiments.
NeuroImage5(4Pt.
1),261–270.
Raz,N.
,Gunning,F.
M.
,Head,D.
,etal.
,1997.
Selectiveagingofthehumancerebralcortexobservedinvivo:differentialvulnerabilityoftheprefrontalgraymatter.
Cereb.
Cortex7(3),268–282.
Spearman,C.
,1904.
Generalintelligenceobjectivelydeterminedandmea-sured.
Am.
J.
Psychol.
15,201–293.
TalairachJ.
,TournouxP.
,1988.
Co-planarstereotaxicatlasofthehumanbrain:a3-dimensionalproportionalsystem,anapproachtocerebralimaging,G.
Thieme,Stuttgart;ThiemeMedicalPublishers,NewYork.
Thompson,P.
M.
,Cannon,T.
D.
,Narr,K.
L.
,etal.
,2001.
Geneticinfluencesonbrainstructure.
Nat.
Neurosci.
4(12),1253–1258.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433432Wechsler,D.
,1981.
WechslerAdultIntelligenceScale—Revised.
Psycho-logicaCorporation,SanAntonio,TX.
White,N.
B.
,Alkire,M.
T.
,Haier,R.
J.
,2003.
Avoxel-basedmorphometricstudyofnon-dementedadultswithDownsyndrome.
NeuroImage20(1),393–403.
Wilke,M.
,Sohn,J.
H.
,Byars,A.
W.
,etal.
,2003.
Brightspots:correlationsofgraymattervolumewithIQinanormalpediatricpopulation.
Neuro-Image20(1),202–215.
Yoshii,F.
,Barker,W.
W.
,Chang,J.
Y.
,etal.
,1988.
Sensitivityofcere-bralglucosemetabolismtoage,gender,brainvolume,brainatrophy,andcerebrovascularriskfactors.
J.
Cereb.
BloodFlowMetab.
8(5),654–661.
R.
J.
Haieretal.
/NeuroImage23(2004)425–433433
- step66smsm.com相关文档
- CS66smsm.com
- related66smsm.com
- Lloyds66smsm.com
- documents66smsm.com
- arriving66smsm.com
- Inspiron66smsm.com
台湾CN2云服务器 2核2G 5M 5IP 台湾物理服务器 E5x2 64G 20M 5IP
提速啦(www.tisula.com)是赣州王成璟网络科技有限公司旗下云服务器品牌,目前拥有在籍员工40人左右,社保在籍员工30人+,是正规的国内拥有IDC ICP ISP CDN 云牌照资质商家,2018-2021年连续4年获得CTG机房顶级金牌代理商荣誉 2021年赣州市于都县创业大赛三等奖,2020年于都电子商务示范企业,2021年于都县电子商务融合推广大使。资源优势介绍:Ceranetwo...
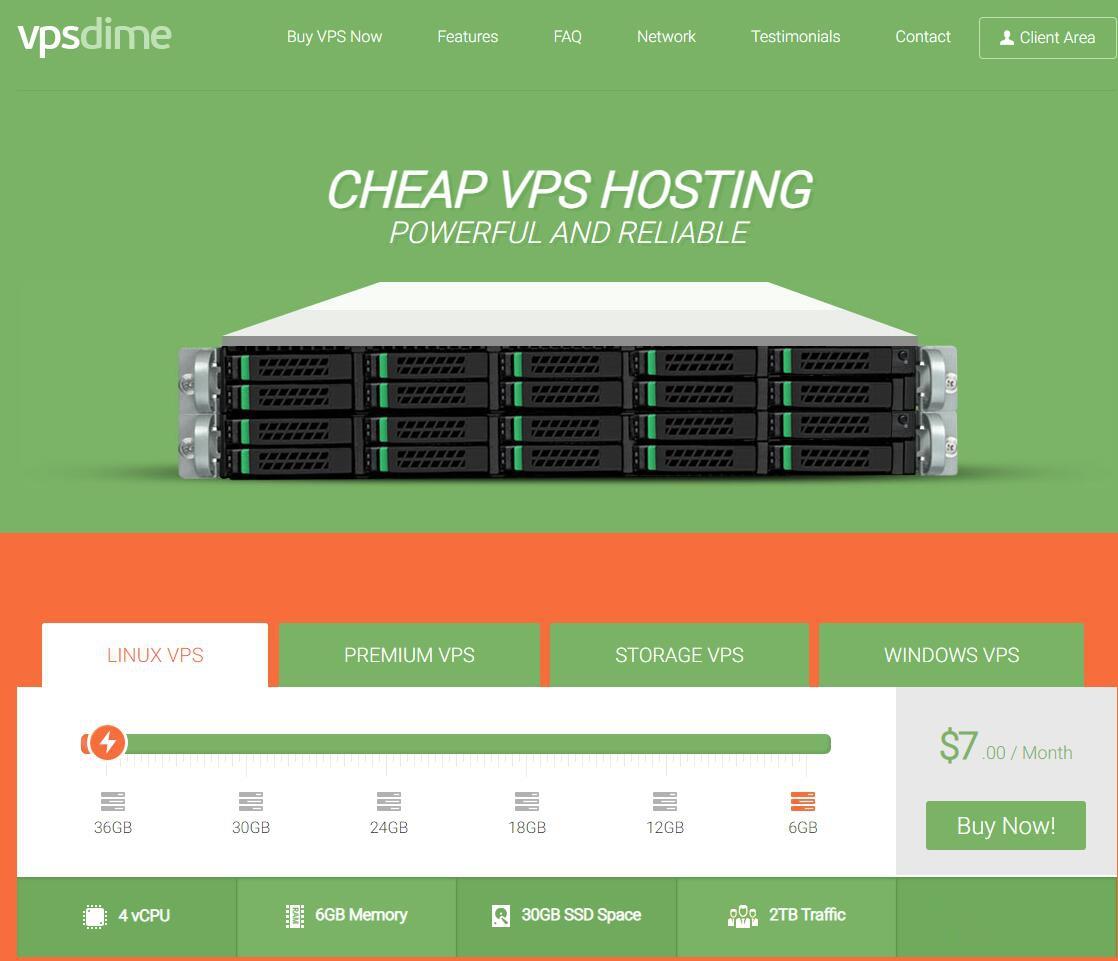
VPSDime7美元/月,美国达拉斯Windows VPS,2核4G/50GB SSD/2TB流量/Hyper-V虚拟化
VPSDime是2013年成立的国外VPS主机商,以大内存闻名业界,主营基于OpenVZ和KVM虚拟化的Linux套餐,大内存、10Gbps大带宽、大硬盘,有美国西雅图、达拉斯、新泽西、英国、荷兰机房可选。在上个月搞了一款达拉斯Linux系统VPS促销,详情查看:VPSDime夏季促销:美国达拉斯VPS/2G内存/2核/20gSSD/1T流量/$20/年,此次推出一款Windows VPS,依然是...
2021年恒创科技618活动:香港/美国服务器/云服务器/高防全场3折抢购
2021年恒创科技618活动香港美国服务器/云服务器/高防全场3折抢购,老客户续费送时长,每日限量秒杀。云服务器每款限量抢购,香港美国独服/高防每款限量5台/天,香港节点是CN2线路还不错。福利一:爆品秒杀 超低价秒杀,秒完即止;福利二:云服务器 火爆机型 3折疯抢;福利三:物理服务器 爆款直降 800元/月起;福利四:DDOS防护 超强防御仅 1750元/月。点击进入:2021年恒创科技618活...

66smsm.com为你推荐
-
汇通物流谁帮我查查百世汇通快递都一天多一直显示发货就是没有物流信息,221202敬汉卿姓名被抢注身份证信息被抢注12306账号怎么办摩根币摩根币到底是什么是不是骗局安徽汽车网安徽省各地车牌号简称是按照什么顺序排的今日油条油条每周最多能吃多少咏春大师被ko练了十几年的 “ 咏春高手”~~被练一年空手道的轻易打败,难道如今的国术就像国足,不堪一击~~75ff.com开机出现www.ami.com是什么?怎么解决啊地陷裂口造成地陷都有哪些原因?杰景新特我准备在网上买杰普特711RBES长笛,10700元,这价格合理吗?还有,这是纯银的吗,是国内组装的吗?www.522av.com我的IE浏览器一打开就是这个网站http://www.522dh.com/?mu怎么改成百度啊 怎么用注册表改啊