Average515hh
515hh com 时间:2021-03-03 阅读:()
RESEARCHOpenAccessGrowthpromotionandcolonizationofswitchgrass(Panicumvirgatum)cv.
AlamobybacterialendophyteBurkholderiaphytofirmansstrainPsJNSeonhwaKim1,ScottLowman1,2,GuichuanHou4,JerzyNowak2,BarryFlinn1,2,3andChuanshengMei1,2,3*AbstractBackground:SwitchgrassisoneofthemostpromisingbioenergycropcandidatesfortheUS.
Itgivesrelativelyhighbiomassyieldandcangrowonmarginallands.
However,itsyieldsvaryfromyeartoyearandfromlocationtolocation.
Thusitisimperativetodevelopalowinputandsustainableswitchgrassfeedstockproductionsystem.
Oneofthemostfeasiblewaystoincreasebiomassyieldsistoharnessbenefitsofmicrobialendophytes.
Results:Wedemonstratethatoneofthemoststudiedplantgrowthpromotingbacterialendophytes,BurkholderiaphytofirmansstrainPsJN,isabletocolonizeandsignificantlypromotegrowthofswitchgrasscv.
Alamounderinvitro,growthchamber,andgreenhouseconditions.
Inseveralinvitroexperiments,theaveragefreshweightofPsJN-inoculatedplantswasapproximately50%higherthannon-inoculatedplants.
Whenone-month-oldseedlingsweregrowninagrowthchamberfor30days,thePsJN-inoculatedAlamoplantshadsignificantlyhighershootandrootbiomasscomparedtocontrols.
Biomassyield(dryweight)averagedfromfiveexperimentswas54.
1%higherintheinoculatedtreatmentcomparedtonon-inoculatedcontrol.
Similarresultswereobtainedingreenhouseexperimentswithtransplantsgrownin4-gallonpotsfortwomonths.
Theinoculatedplantsexhibitedmoreearlytillersandpersistentgrowthvigorwith48.
6%higherbiomassthancontrols.
WealsofoundthatPsJNcouldsignificantlypromotegrowthofswitchgrasscv.
Alamoundersub-optimalconditions.
However,PsJN-mediatedgrowthpromotioninswitchgrassisgenotypespecific.
Conclusions:OurresultsshowB.
phytofirmansstrainPsJNsignificantlypromotesgrowthofswitchgrasscv.
Alamounderdifferentconditions,especiallyintheearlygrowthstagesleadingtoenhancedproductionoftillers.
Thisphenomenonmaybenefitswitchgrassestablishmentinthefirstyear.
Moreover,PsJNsignificantlystimulatedgrowthofswitchgrasscv.
Alamoundersub-optimalconditions,indicatingthattheuseofthebeneficialbacterialendophytesmayboostswitchgrassgrowthonmarginallandsandsignificantlycontributetothedevelopmentofalowinputandsustainablefeedstockproductionsystem.
Keywords:Bacterialendophyte,BurkholderiaphytofirmansstrainPsJN,Colonization,Growthpromotion,Biomassincrease,Switchgrasscv.
Alamo*Correspondence:chuansheng.
mei@ialr.
org1InstituteforSustainableandRenewableResource,InstituteforAdvancedLearningandResearch,150SlaytonAve,Danville,VA24540,USA2DepartmentofHorticulture,VirginiaPolytechnicInstituteandStateUniversity,Blacksburg,VA24601,USAFulllistofauthorinformationisavailableattheendofthearticle2012Kimetal.
;licenseeBioMedCentralLtd.
ThisisanOpenAccessarticledistributedunderthetermsoftheCreativeCommonsAttributionLicense(http://creativecommons.
org/licenses/by/2.
0),whichpermitsunrestricteduse,distribution,andreproductioninanymedium,providedtheoriginalworkisproperlycited.
Kimetal.
BiotechnologyforBiofuels2012,5:37http://www.
biotechnologyforbiofuels.
com/content/5/1/37BackgroundIncreasingconcernoverforeignenergysupplies,globalgreenhousegasemissionsandtheneedforruraleco-nomicdevelopmenthasdriventheinterestinsustain-ablebiomassproductionforbioenergyandbio-products.
Ithasbeensuggestedthatby2025,theworldenergyde-mandwilllikelybeincreasedbymorethan50%[1,2].
Thisdemand,andsocietalconcernsabouttheenviron-mentalimpactofburningfossilfuelsarekeyfactorsstimulatingthedevelopmentofnationalandregionalstrategiesaimedatthegrowthofrenewableenergysup-plies,primarilyfocusedonbiofuels.
Toreducethereli-anceonfossilfuels,theUSA,theworld'smajorenergyconsumer,releasedtheEnergyIndependenceandSecur-ityActof2007thataimstoincreasetheproductionofrenewablefuelsfrom9.
0billiongallonsin2008to36billiongallonsby2022[3].
TherecentUSDA/DOENa-tionalBiofuelsActionPlan[4]hashelpedtodelineatethepriorityareasrequiredtoacceleratesustainablebio-fuelindustrydevelopment.
Withinthisdocument,Ac-tionArea2wasidentifiedasfeedstockproductionandimprovement.
Variousfeedstocks,suchasperennialrhi-zomatousgrasses,canprovidesourcesoflignocellulosicbiomass,servingasnewsourcesofcropgrowthandin-comeforregionalfarmers.
Oneofthemostpromisingfeedstockscapableofcon-tributingtotherealizationofUSrenewableenergygoalsisthecommonperennialgrass,switchgrass(PanicumvirgatumL.
)[5].
Thisnativeprairiegrass,consistingofadiversegermplasm[6],cangrowonmarginallandsunderlowinputsofwaterandagrochemicals[7],sothatitscultivationdoesnotcompetewithfoodcropsforlandandotherresources.
Duetoitslargerootsystemandfaststandregrowth,switchgrasshasotherpositiveenvir-onmentaleffects,includingthepreventionofsurfacerunoffandsoilerosion,carbonsequestration,andtheprovisionofawildlifehabitat[5,8].
Switchgrassculti-vatedlandsalsohadmuchhighertotalsoilorganiccar-bondepositsthanlandscultivatedwithannualcrops,suchascornandwheat[9,10].
Theeconomicsofbiofuelproductionishighlydependentonfeedstockcostandconversiontechnology[1,6].
Thedevelopmentofimprovedswitchgrassvarietiesforlow-costproductiononmarginallandsisoneprerequisiteforthesuccessofthebioenergyprogram[5,11].
Onesuchap-proachinvolvestheuseofbeneficialmicroorganisms,suchasendophytes,whichformintimateassociationswithplants[12,13].
Endophytes,bothfungalandbacterial,havebeentargetedasmechanismstoenhanceplantcharacteris-ticsforcommercialuses[14].
Thecolonizationofgrassesbyfungalendophytesforperformanceenhancementiswelldocumented[15],includingtheirusewithswitchgrass[16,17].
However,toourknowledge,onlyonestudyhasreportedgrowthpromotionofabioenergyfeedstockgrass(Miscanthusxgiganteous)seedlingsbyabacterialendophyte(Herbaspirillumfrisingense)[18].
Akeycom-ponentofourbioenergycropresearchprograminvolvestheutilizationofbeneficialbacterialendophytesthatformstableandpersistentassociationswithswitchgrass,asthemechanismtoimprovebiomassyieldandenhancestresstoleranceunderlow-inputproductionsystems[19].
Bene-ficialbacterialendophytesarenaturallyoccurringsoilmicroorganismsthatcanpenetrateplantrootsandtrans-locatetotheabovegroundorgansand,uponcolonization,affectplantgrowth,health,andproductivity[12,20-22].
Althoughthemolecularmechanismsofbeneficialendophyte-hostplantinteractionsarelargelyunknown,severalstudieshavedemonstratedthatendophytescanpromoteplantgrowthbyenhancingtheplant'scapacityfornutrientacquisition,betterwatermanagement,and/orresistancetoabioticandbioticstressesviaregulationofhormones[12-14,20,21].
Forinstance,1-aminocyclo-propane-1-carboxylicacid(ACC)deaminaseproducedbyendophyteslowerstheethylenelevelsinhostplants,redu-cingtheirresponsetoabioticandbioticstress,andbychangingrootmorphology,leadingtostimulationofplantgrowth[13,23,24].
Manyknownendophytesalsopromoteplantgrowthbyproducinggibberellicacid(GA3),indole-3-aceticacid(IAA)[18,25],orcytokinins[26,27].
BurkholderiaphytofirmansstrainPsJNhasbeenfoundtobeahighlyeffectiveplantgrowthpromotingbacterialendophyte,withabroadhostrangeincludingpotatoes,tomatoes,andgrapevines[21,27-32].
Inaddition,itsgenomehasrecentlybeensequenced[33],providingthegenomicresourcesneededtodevelopanunderstandingofthemechanismsassociatedwiththisendophyte'sabilitytopromoteplantgrowth.
PsJNpro-ducesahighlevelofACCdeaminase[31],enhanceshostplantcold[34]andheat[35]stresstolerance,improveswatermanagement[36]andplantresistancetopathogens[37,38].
Inthisstudy,wereportgrowthpromotionofswitchgrasscv.
AlamobyBurkholderiaphytofirmansstrainPsJNunderinvitro,growthcham-ber,andgreenhouseconditions.
Toourknowledge,thisisthefirstreportdetailingtheswitchgrass-PsJNinteraction.
ResultsPsJNendophyticassociationwithswitchgrassAlamoTheendophyticcolonizationofswitchgrassbyBurkhol-deriaphytofirmansstrainPsJN-GFPwasvisualizedusingconfocalmicroscopy(Figure1).
Undertheappropriateillumination,thePsJN-GFPcouldbeclearlyobservedin-sidetherootsofPsJN-inoculatedplants3daysafterin-oculation,whilenofluorescencewasobservedinrootsofbuffer-inoculatedcontrolplants.
ThetiterofPsJN-GFPininoculatedplantswasalsodeterminedusingtis-suehomogenatesfromvarioustissues(root,leafandKimetal.
BiotechnologyforBiofuels2012,5:37Page2of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37sheath)atdifferenttimes(Table1).
Theendophyteini-tiallyinfectedandcolonizedplantroots,andby7dayspost-inoculation,PsJNtiterswerestillhighestintheroot.
However,thetitersincreasedsignificantlyinothertissuesbyday14,indicatingtranslocationtoleavesandsheaths.
EffectsofPsJNonAlamogrowthinvitroYoungswitchgrassseedlingswerepreparedandinocu-latedasdescribedinMaterialsandMethods,andthenon-inoculatedandinoculatedplantswereanalyzedaftergrowthinvitroforonemonth.
TheresultshowedthatPsJNsignificantlyandrepeatedlypromotedAlamorootandshootgrowth,witha35.
6%increaseinshootlength,a32.
8%increaseinrootlength,andan83.
6%in-creaseinfreshweight,comparedtothenon-inoculatedplants(Figure2).
Afterseveralreplications,theaveragefreshweightofthePsJN-inoculatedplantswasalwaysapproximately50%higherthannon-inoculatedplants.
EffectsofPsJNonAlamogrowthinagrowthchamberenvironmentAsdescribedabove,PsJNsignificantlyenhancedswitch-grasscv.
Alamogrowthinvitro.
Wenextassessedtheim-pactofPsJNongrowthundersoilconditions.
One-month-oldinvitrogrownAlamo(controlandPsJN-inoculatedseedlings)weretransferredtoaflatwith72cavitiesfilledwithsoilandgrowninagrowthchamberunder28/22°Cday/nighttemperatureswith16hlightperiodforonemonth.
ThePsJN-inoculatedplantsshowedsignificantgrowthincreasescomparedtocontrolplantsinshootlengthandfresh/dryweights(Figure3).
Thegrowthcham-berexperimentswererepeated5times,andtheaveragedatafrom5experimentsshowedsignificantgrowthpro-motionbyPsJN,witha46.
3%,anda54.
1%increaseinfreshweightanddryweight,respectively.
Thetotaldryweightincrease(54.
1%)byPsJNwasmorethanthetotalfreshweightincrease(46.
3%),indicatingthatthePsJN-inoculatedplantsaccumulatedmorebiomass.
EffectsofPsJNonAlamogrowthinthegreenhouseNon-inoculatedandPsJN-inoculatedplantswerealsogrownundergreenhouseconditionstodeterminegrowthenhancementpersistence.
TheplantsinoculatedwithPsJNandgrowninvitrofor25daysweretransferredto4-gallonpotswith5plantsineachpotandgrowninthegreen-house.
TheplantsinoculatedwithPsJNexhibitedsustainedgrowthvigor,astheyweresignificantlytaller,andmoretil-lersdevelopedearlycomparedwiththenon-inoculatedcontrolplants(Figure4).
Followingonemonthofgrowthinthegreenhouse,thePsJN-inoculatedplantshad76.
2%moretillersthanthecontrolplants.
TheplantswereFigure1Confocalimagesofrootstaken3daysfollowingswitchgrasscv.
AlamoinoculationwithPsJN-GFP,showingbacterialcolonizationinsidetheroots.
(A):Controland(B):PsJN-inoculatedplants.
(C):Controland(D):PsJN-inoculatedplants.
Ineachsection(a,b,c,d),leftpanelswereobservedunderfluorescentlightandrightpanelsundervisiblelight.
Thebarsrepresent100μm(AandB)and20μm(CandD).
Table1Colony-formingunits(CFU)ofroot,leaf,andsheathtissuesatdifferenttimesafterPsJN-GFPinoculationDaysafterPsJN-PlanttissuesAverageCFU/gGFPinoculationFreshweight3All(Roots,leaves,andsheath)4.
2X1057Roots7.
6X105Leaves2.
6X10314Roots3X104Sheaths1.
3X105Leaves1.
2X105Kimetal.
BiotechnologyforBiofuels2012,5:37Page3of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37harvestedfollowinggrowthfortwomonths,andthebio-massyielddetermined(Figure5).
ThePsJN-inoculatedplantswererepeatedlysignificantlyhigherinbiomassyield,witha36.
8%increaseinfreshweightanda57.
1%increaseindryweight.
EffectsofPsJNonAlamogrowthinsub-optimalconditionsInordertodevelopalowinputandsustainableswitch-grassfeedstockproductionsystemutilizingthebeneficialbacterialendophyte,wetestedgrowthperformanceofPsJN-inoculatedplantwithunfertilizedfieldsoil,inaglasshouseunderambientconditionsduringtheFall,whenthetemperaturewasnotoptimalforswitchgrassgrowth.
TheresultsshowedthatPsJN-inoculatedplantsproducedtwicethetotalbiomassofcontrols(Figure6).
DirectinoculationofswitchgrassseedswithPsJNInordertoexploreapracticalwaytoinoculateswitch-grasswiththebacterialendophyte,westerilizedswitch-grassseedsasdescribedinMaterialsandMethods,placedthesterilizedseedsonwetfilterpaperfor3-5daysinanincubatorat25°C,andtheninoculatedthegerminatingseedswithdifferentconcentrationsofendophyteinocu-lumtodeterminetheoptimalinoculationconcentration(OD600at0.
1-0.
5).
TheplantsinoculatedwithPsJNatOD600of0.
1,0.
25,and0.
5exhibited28.
7%,55.
0%and80.
1%increasesindryweight,respectively,comparedtonon-inoculatedplantsaftergrowninvitrofor25daysandingrowthchamberforanothermonth.
APsJNconcentra-tionof0.
5wasthemosteffectiveatpromotingbiomassincrease(Figure7),andnobiomassdifferencewasobservedbetweenthe0.
1treatmentandcontrol.
Endophyteinfectionandcolonizationofseedsaredependentonendophyteconcentrationandthestatusofseedimbibitions.
So,inordertofacilitateinfectionandcolonizationbythebacterialendophyte,thesterilizedseedswereimbibedinwaterfor1,2,3,or4days,andtheninoculatedwithPsJNatanOD600of0.
5or1.
0,sinceanOD600of0.
5wasthemosteffectiveasdescribedabove.
ThePsJN-inoculatedseedswereplacedinanin-cubatorat25°Cwitha16hlightperiodfor25days,andtransferredtosoilandgrowninagrowthchamberfor37days.
Theresultsindicatedthatplantsfromtheseedsimbibedfor2daysandtheninoculatedwithanOD600of0.
5hadthehighestdryweight,witha55%increasecomparedtonon-inoculatedcontrolplants(Figure8).
GenotypicresponsestoPsJNAsdescribedabove,PsJNwasabletostimulategrowthinswitchgrasscv.
Alamo.
ToassesstheinfluenceofFigure2EffectsofendophytePsJNinoculationonswitchgrasscv.
Alamogrowthinvitro.
Datawereobtainedafterplantsweregrowninincubatorfor36days.
Samplenumberwas25,and**meanssignificantdifferenceat0.
01levelbetweenPsJNandcontrolusingstudentT-test.
Figure3EffectsofendophytePsJNinoculationonswitchgrasscv.
Alamogrowthingrowthchamber.
TheseedlingswereinoculatedwithPsJNandgrowninvitroforonemonth,thentransferredtosoilandgrowningrowthchamberforanothermonth.
Dryweightwasdeterminedaftersamplesweredriedinovenat65°Cforoneday.
Samplenumberwas36,and**meanssignificantdifferenceat0.
01levelbetweenPsJNandcontrolusingstudentT-test.
Kimetal.
BiotechnologyforBiofuels2012,5:37Page4of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37plantgenotypeonthisresponse,sevenotherswitchgrasscultivarsweretestedfortheirgrowthresponsestoPsJN.
AsshowninTable2,growthpromotionbyPsJNwasgenotype-dependent.
Theswitchgrasscvs.
Forestburg,Nebraska,andBlackwellwereallresponsivetoPsJN,withmeasuredsignificantgrowthincreasesof60.
1%,26.
8%,and23.
0%,respectively,whilethecvs.
Cave-in-Rock,Sunburst,Shelton,andShawneedidnotexhibitgrowthpromotioninresponsetoPsJNundersimilarconditions.
PreliminaryresultfromCave-in-Rockbio-assayindicatedthatPsJNtiterswerenotsustainedafterinoculationandweremuchlowerinthenon-responsiveplantsfollowinginoculation.
DiscussionInthepresentstudy,wedemonstratedtheabilityofB.
phytofirmansPsJNtocolonizeandpromotegrowthinswitchgrasscv.
Alamo.
ThreedaysfollowingPsJNinocu-lation,wecouldclearlyvisualizebacterialcolonizationinsidetherootsunderconfocalmicroscopy.
Thebacter-ialpopulationinsidetherootswasinitiallymuchhigherthanthatoftheleavesandsheaths,andthebacterialendophytewassubsequentlytranslocatedverticallytotheupperleavesthroughtheleafsheath.
Theseresultsweresimilartothatreportedforgrapevine[22]andpo-tato[39],wherePsJNwastransportedthroughtheinter-iorvascularsystem,fromrootxylemvesselstotheupperpartsoftheplants.
Thisisacriticalfirststepintheendophyticbacteria-plantinteraction[40].
Weobservedsignificantgrowthpromotionofcv.
AlamobyPsJN,underbothinvitroandsoilconditions.
Ourstudyshowedtotalfreshweightandtotaldryweightoftheinoculatedplantswasincreasedby45%and55%respect-ivelycomparedwiththenon-inoculatedcontrolplantswhentheinoculatedseedlingsweregrowninvitroandthentransferredtosoilandgrowningrowthchamberforonemonth.
Similarresultshavebeenobtainedin4-gallonpotsunderourgreenhouseconditions.
OtherstudieshavereportedlevelsofgrowthpromotionbyPsJN,withgrapevineshowinga6-foldincreaseintotalbiomass[34],andpotatoshowinganapproximate2-foldincreaseinrootandhaulmbiomass[36]overcontrols.
ThemechanismofplantgrowthbyB.
phytofirmansPsJNhasbeenreported[21]andattributedtotheabilityFigure4PlantgrowthandtillerdevelopmentaftercontrolandPsJNinoculatedplantsweretransferredto4-gallonpotsandgrowningreenhouse.
Figure5GrowthpromotionpersistenceofAlamoingreenhousebyPsJNinoculation.
TheseedlingswereinoculatedwithPsJNandgrowninvitroforonemonth,thentransferredto4-gallonpotwith5plants/potandgrowningreenhousefortwomonths.
Dryweightwasdeterminedaftersamplesweredriedinovenat65°Cforoneday.
Samplenumberwas25,and**meanssignificantdifferenceat0.
01levelbetweenPsJNandcontrolusingstudentT-test.
Kimetal.
BiotechnologyforBiofuels2012,5:37Page5of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37ofPsJNtoproducehighlevelsofACCdeaminaseactiv-ity,whichdegradesACCtoammoniaandα-ketobutyrate[41],whichisacommoncharacteristicofplantgrowthpromotingbacteria.
ACCistheprecursortoethylene,aplantstresshormone;hence,thereducedethylenelevelinPsJN-colonizedplantswillpromoteplantgrowth.
AccordingtothereportbyPenroseandGlick(2003)[42],ACCactivityover20nmolα-ketobutyrate/h/mgissufficienttopromotehostplantgrowth,andPsJNhasbeenshowntocontain308nmolα-ketobutyrate/h/mgofACCdeaminaseactivity[31].
Al-thoughseveralstudieshavereportedtheinteractionbe-tweenthisendophyteandhostplantsforgrowthpromotion,moststudieshavereportedinvitrodata[22,27,28,36].
Ourresultswithunfertilizedfieldsoil,inaglasshouseunderambientconditionsduringtheFall,whenthetemperaturewasnotoptimalforswitchgrassgrowth(Figure6)impliedthepotentialbenefitofswitch-grassinoculatedwithPsJNforgrowthonmarginallandsandsub-optimalgrowthconditions.
Althoughourresultsindicatepotentialbenefitofbacterialendophytesinswitchgrassunderinvitro,growthchamberandgreen-houseconditions,muchworkstillneedstobedoneinassessinggrowthpromotionofswitchgrassbyendo-phytesinthefieldsincethereismuchmorecompetitionbetweenendophytesandothermicroorganismsinadditiontomanyotherunfavorableconditions.
Whileourinitialstudieswerecarriedoutwithcv.
Alamo,wetestedtheutilityofPsJNasagrowth-promotingendo-phytewithotherswitchgrasscultivars.
Ourresultsindi-catedthatspecificgenotypeeffectsexisted,withsomegenotypesbeinghighlyresponsivetothegrowthpromo-tiveeffectsofPsJN,andothersnot.
Similargenotypeeffectshavebeenreportedbyothers.
ItwasreportedthatthepotatoresponsetoPsJNinvolvessomeformofgen-eticcontrol,assomepotatocultivarsdisplaythebenefi-cialresponsetotheendophyte,whileothersdonot[35,43,44].
Thetypicalinvitrophenotypeforastronglyresponsivecultivarwascharacterizedbyamassive,well-branchedrootsystemandafterthefirst3-4weeksinFigure6GrowthpromotionofAlamobyPsJNinoculationinsub-optimalconditions.
TheseedlingswereinoculatedwithPsJNandgrowninvitroforonemonth,thentransferredto4-gallonpotswith5plants/potwithunfertilizedfieldsoilandgrownintheglasshouseunderambientconditionsfor2.
5monthsinthelateFallof2010.
Dryweightwasdeterminedaftersamplesweredriedinovenat65°Cforoneday.
Samplenumberwas25,and**meanssignificantdifferenceat0.
01levelbetweenPsJNandcontrolusingstudentT-test.
Figure7EffectsofdifferentconcentrationsofPsJNonswtchgrasscv.
Alamogrowthwithdirectseedinoculation.
Thesurface-sterilizedseedswereinfectedwithdifferentconcentrationsofPsJNandgrowninvitrofor17days,thentransferredto72-cavitytraysandgrowninagrowthchamberfor50days.
*and**meansignificantdifferenceat0.
05and0.
01levelsrespectivelybetweenPsJNandcontrolusingstudentT-test.
Kimetal.
BiotechnologyforBiofuels2012,5:37Page6of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37culture,theplantletwasdevelopmentallymoreadvancedthanthenon-bacterizedcontrols.
Stemsweresturdier,withmorelignindepositsaroundthevascularsystem,andtheplantletsdevelopedmoreroothairsandmoreandlargerleaftrichomes[43].
WealsonoticedPsJN-inoculatedswitchgrassplantsweredevelopmentallyadvanced(unpublisheddata).
Suchenhancementswerenotapparentforthepoorly-responsivecultivars.
WealsoobservedsomeofthesephenotypicdifferencesbetweenPsJN-responsiveandnon-responsiveswitchgrassculti-vars.
AdditionalworkillustratingthegeneticcontrolofthebeneficialresponsetotheendophyteusedmonoploidpotatolinesderivedfromanthercultureofanadapteddiploidSolanumphurejaclone,BARD1-3[44].
Thedip-loidantherdonor,BARD1-3,exhibitedabacterizationresponsecomparabletoRedPontiac,whilemonoploidlinesexhibitedaresponsetoPsJNrangingfromfavorabletounfavorabletoneutral.
Theassumptionherewasthattheresponserangeofthemonoploidpopulationwasduetothesegregationofallelesforgenesinvolvedinregulat-ingthepositiveornegativeinteractionwithPsJN.
Thepotato/PsJNstudieshavebeenthemostcharacter-ized,andcarriedoutwithmaterialclonallypropagatedvianodalsections,inwhichasingleinoculationissuffi-cienttoinitiatecolonizationthroughthexylemtissue,Figure8EffectsofdifferentconcentrationsofPsJNonswitchgrasscv.
Alamogrowthwithdirectinoculationofseedsthatwereimbibedinwaterfordifferentdays(1-4days).
TheseedswereinfectedwithdifferentconcentrationsofPsJNandgrowninvitrofor25days,thentransferredto72-cavitytraysandgrowninagrowthchamberfor37days.
Samplenumberwas72foreachtreatment.
*and**meansignificantdifferenceat0.
05and0.
01levelsrespectivelybetweenPsJNandcontrolusingstudentT-test.
Table2EffectsofB.
phytofirmansPsJNonplantgrowthindifferentswitchgrasscultivarsCultivarsTreatmentNo.
plantsRootlength(cm)Shootlength(cm)Totalfreshweight(mg)PsJN/control(%)aShawneeControl242.
38.
450.
1PsJN242.
17.
858.
8117.
4p-valueb0.
20370.
20490.
0717NebraskaControl241.
510.
340.
9PsJN241.
511.
651.
9126.
8p-value0.
41600.
06560.
0055ForestburgControl341.
68.
633.
7PsJN301.
812.
754.
0160.
1p-value0.
19860.
00000020.
0000001SheltonControl283.
314.
1117.
7PsJN283.
012.
0135.
3115.
0p-value0.
15790.
00670.
0907BlackwellControl281.
49.
952.
6PsJN281.
511.
364.
7123.
0p-value0.
29700.
09980.
0543SunburstControl300.
88.
533.
0PsJN280.
910.
426.
480.
0p-value0.
29850.
01370.
0731Cave-in-RockControl332.
814.
7107.
8PsJN343.
113.
8113.
4105.
2p-value0.
23170.
40520.
5613aPsJN/control(%)meanstotalfreshweightofPsJNinoculatedplantsoverthatofcontrolplants.
bp-valuemeansdegreesofstatisticalsignificancebetweenPsJNtreatmentandcontrolusingstudentT-test.
Kimetal.
BiotechnologyforBiofuels2012,5:37Page7of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37eventuallyspreadingtotheupperleaves[36].
Bacteriallevelsmustreachathresholdpopulationwithintheplantbeforetheyareeffective[45]withadirectrelation-shipbetweenplantletgrowthenhancementandPsJNcolonizationofbothinteriorandexteriorsurfaces[44].
ThePsJNcolonizationprofilesforaresponsiveandpoorlyresponsivecultivarover8tissueculturegenera-tionsrevealedbacterialloadsoneorderofmagnitudegreaterforshoot/rootsurfaceandinteriorcolonizationintheresponsivecomparedtothepoorlyresponsivecul-tivar.
Furthermore,theresponsivecultivarexhibitedincreasedcolonizationoversuccessivegenerations,whilethepoorlyresponsivecultivarexhibiteddecliningbacter-ialpopulationsoversuccessivegenerations.
Wearecur-rentlyassessingthelevelofcolonizationinswitchgrasscultivarsresponsiveandnon-responsivetoPsJNtode-terminethedegreeofsimilaritybetweenswitchgrassandpotatoresponsestotheendophyte.
Atpresent,themechanismsgoverningB.
phytofirmansPsJNgenotype-specificityingrowthpromotionofswitchgrass(andotherplants)areunknown,althoughwearecurrentlyusinglargescalegeneexpressionanalysestodeterminethedifferencesintheswitchgrassmolecularresponsesbetweendifferently-respondingcultivars.
Insummary,theresultsreportedhereillustratetheabilityofB.
phyto-firmansPsJNtoinfectandcolonizeresponsiveswitch-grass(Panicumvirgatum)cultivarslikeAlamo,andtopromoteplantgrowth.
Thisstudylaysthefoundationtodevelopalowinputandsustainableswitchgrassfeed-stockproductionsystemonmarginallandsusingthis,andother,beneficialbacterialendophytes.
ConclusionsResultsobtainedwithgrowthpromotionofswitchgrasscv.
AlamobyB.
phytofirmansPsJNundervariousgrowthconditions,sub-optimalinparticular,indicatethepotentialforutilizationofbeneficialbacterialendo-phyteinswitchgrassestablishmentinthefirstyearandinthedevelopmentoflowinputandsustainableswitch-grassfeedstockproductionsystem.
Inthefuture,themechanismsofgrowthpromotionneedtobeelucidatedwithmolecularbiologyandfunctionalgenomicstode-veloptoolsformolecularbreedingforbeneficialplant-microbialassociations.
MethodsPlantmaterialsSwitchgrass(PanicumvirgatumL.
)cvs.
AlamoandCave-in-RockseedswerepurchasedfromWarnerBroth-ersSeedCo.
(Lawton,OK),andotherswitchgrassseedswerekindlyprovidedbyDr.
BingyuZhao(DepartmentofHorticulture-VirginiaTech,Blacksburg,VA).
Switch-grassseedsweresurface-sterilizedbytreatmentwith70%ethanolfor2min,rinsed3Xwithdistilledwater,de-huskedfor30minwith60%H2SO4withstirring,washed3Xwithdistilledwater,sterilizedwith0.
4Mso-diumhypochlorite(50%commercialbleachsolutioncontaining6%sodiumhypochlorite)containing0.
1%Triton100for30minfollowedby5Xrinsewithsterile,deionized,distilledwater(ddH2O).
BacterialendophyteandcultureconditionsBurkholderiaphytofirmansstrainPsJN[31]anditsPsJN-GFPderivative[22]wereobtainedfromDr.
AngelaSessitsch(AustrianInstituteofTechnology,Seibersdorf,Austria).
ThecultureswerestreakedonKing'sB(KB)solidmediumasdescribedin[45].
Inoculumwaspro-ducedbytransferringoneloopofPsJNfrom2-day-oldculturesto5mlKBbrothina15-mlculturetube,fol-lowedbyincubationat28°Conashaker(150rpm)over-night.
FivemloftheovernightPsJNculturewasaddedto45mlKBbrothina250-mlErlenmeyerflaskandgrownto0.
7OD600.
Bacterialcellswerethencollectedbycentrifugationat3,500rpmfor7minat4°C,andre-suspendedinPBSbuffer(10mMNaH2PO4containing0.
8%NaC1,pH6.
5)afterwhichtheOD600wasadjustedwithPBSbufferto0.
5,unlessdescribedotherwise.
SeedlinginoculationwithPsJNandplantgrowthresponsesSurface-sterilizedseedsweregerminatedinpetri-dishesfor7daysat25°C,underwhitefluorescentlight(67μmolm-2s-1),16hphotoperiod,onaswitchgrassgrowthmediumconsistingofMSsaltsandvitamins[46],30g/lmaltoseand3g/lphytogel,pH5.
8.
TheroottipsoftheyoungseedlingswerecutpriortoPsJNinocu-lationtofacilitatebacterialpenetration[45].
Forthedir-ectseedinoculationsurface-sterilizedseedswereplacedonwetfilterpaperfor3-5daysinanincubatorat25°Cwith16hphotoperiod(whitefluorescentbulbsat67μmolm-2s-1)followedbysoakinginPsJNsolution(0.
5ofOD600)for1min.
Controlseedlings/seedsweretreatedwithPBSbufferalone.
Thetreatedseedling/seedswereblot-driedwithsterilepapertowel,placedonswitchgrassgrowthmediuminGA7Magentavessels(Sigma-Aldrich)containing50mlofmediaand5seed-lingsorgerminatingseedspervessel,andgrownforonemonthintheincubatorasabove.
Rootandshootlength,andseedlingfreshweightwerethendetermined,andtheplantstransferredtoasoilmixcomposedof2/3Miracle-GroWPottingMix(ScottsMiracle-GroCom-pany,Marysville,Ohio)and1/3Arabidopsisgrowingmedia(LehleSeeds,RoundRock,Texas).
Plantsweregrownin72-cavitytraysinagrowthchamberata28/22°Cday/nighttemperature,16hphotoperiod(whitefluorescentbulbsat88μmolm-2s-1)for30days,or4-gallonpotsinthegreenhouse.
Kimetal.
BiotechnologyforBiofuels2012,5:37Page8of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37PsJNcolonizationTheplantsinoculatedwithPsJN-GFPwereexaminedunderafluorescentstereomicroscope(ModelSZX-ILLD2-100;Olympus,Tokyo,Japan)equippedwithaGFPfilter(BP460-490,Olympus,Tokyo,Japan)andtheZeiss510laserscanningconfocalmicroscope(LSCM)(CarlZeiss,Inc.
,Thornwood,NY)toobservecolonization.
Forbioassays,thecontrolandPsJN-GFPinoculatedplantsweresurface-sterilizedwith0.
032Msodiumhypo-chloritefor1min,thenwashed4Xwithsteriledistilledwater.
FiftyμlofthefinalwashwasplatedonsolidKBmediumtoconfirmeffectivenessofsurfacesterilization.
Root,leafandsheathpartswerethenseparated,eachweighed,andgroundwithmortarandpestlein1mlster-iledistilledwater.
Thehomogenateswerethencentrifugedat2000rpmfor3min,andthesupernatantdilutedto1:10,1:100,and1:1000withdistilledwater.
Fiftyμlsam-plesoftheseriallydilutedsolutionswerespreadonsolidKBmedium.
Theplateswereincubatedfor3daysat28°CinthedarkandthenumberofGFP-positivecoloniesdeterminedusingfluorescencestereomicroscopyasdescribedabove.
CompetinginterestsTheauthorsdeclarethattheyhavenocompetinginterests.
Authors'contributionsSKandSLconductedexperimentsanddraftedthemanuscript.
GHcarriedoutconfocalimaginganddataanalysisandreviewedthemanuscript.
CM,JNandBFdesignedexperimentsandwrotethepaperwithinputfromallauthors.
Allauthorsreadandapprovedthefinalmanuscript.
AcknowledgementsThisworkwasfundedthroughSpecialGrants(2003–38891–02112,2008-38891-19353and2009-38891-20092)andHATCHfunds(ProjectNo.
VA-135816)fromtheUnitedStatesDepartmentofAgriculture,theOfficeofScience(BER),U.
S.
DepartmentofEnergyforPlantFeedstockGenomicsforBioenergyProgram(DE-SC0004951),andoperatingfundsfromtheCommonwealthofVirginiatotheInstituteforAdvancedLearningandResearch.
Authordetails1InstituteforSustainableandRenewableResource,InstituteforAdvancedLearningandResearch,150SlaytonAve,Danville,VA24540,USA.
2DepartmentofHorticulture,VirginiaPolytechnicInstituteandStateUniversity,Blacksburg,VA24601,USA.
3DepartmentofForestResourcesandEnvironmentalConservation,VirginiaPolytechnicInstituteandStateUniversity,Blacksburg,VA24601,USA.
4TheDewelMicroscopyFacilityattheCollegeofArtsandSciences,AppalachianStateUniversity,Boone,NC28608,USA.
Received:26March2012Accepted:10May2012Published:30May2012References1.
HamelinckCN,HooijdonkDV,FaaijAP:Ethanolfromlignocellulosicbiomass:techno-economicperformanceinshort-,middle-,andlong-term.
BiomassBioenergy2005,28:384–410.
2.
ErahinME,GomecCY,DereliRK,ArikanO,OzturkI:Biomethaneproductionasanalternativebioenergysourcefromcodigesterstreatingmunicipalsludgeandorganicfractionofmunicipalsolidwastes.
JBiomedBiotechnol2011,201(1):951043.
doi:10.
1155/2011/951043.
3.
SissineF:EnergyIndependenceandSecurityAct.
;2007[http://www.
energy.
senate.
gov/public/index.
cfm/files/serveFile_id=82061c68-eef5-4fc3-81b6-ecd569a1d8a3].
4.
NationalBiofuelsActionPlan:2008[http://www1.
eere.
energy.
gov/biomass/pdfs/nbap.
pdf].
5.
SandersonM,AdlerPR,BoatengAA,CaslerMD,SarathG:SwitchgrassasabiofuelsfeedstockintheUSA.
CanJPlantSci2006,86:1315–1325.
6.
McLaughlinSB,KszosLA:Developmentofswitchgrass(Panicumvirgatum)asabioenergyfeedstockintheUnitedStates.
BiomassBioenergy2005,28:515–535.
7.
HillJ,NelsonE,TilmanD,PolaskyS,TiffanyD:Environmental,economic,andenergeticcostsandbenefitsofbiodieselandethanolbiofuels.
ProcNatlAcadSciUSA2006,103:11206–11210.
8.
HumphreysMO:Thecontributionofconventionalplantbreedingtoforagecropimprovement.
Saskatoon:Proceedingsofthe18thInternationalGrasslandCongress;1999.
9.
LiebigMA,JohnsonHA,HansonJD,FrankAB:Soilcarbonunderswitchgrassstandsandcultivatedcropland.
BiomassBioenergy2005,28:347–354.
10.
McLaughlinSB,UgarteDGLT,GartenCT,LyndLR,SandersonMA,TolbertVR,WolfDD:High-valuerenewableenergyfromprairiegrasses.
EnvironSciTechnol2002,36:2122–2129.
11.
DyerJM,MullenRT:Engineeringplantoilsashigh-valueindustrialfeedstocksforbiorefining:theneedforunderpinningcellbiologyresearch.
PhysiolPlant2008,132:11–22.
12.
SturzAV,ChristieB,NowakJ:Bacterialendophytes:potentialroleindevelopingsustainablesystemofcropproduction.
CritRevPlantSci2000,19:1–30.
13.
BergG:Plant-microinteractionspromotingplantgrowthandhealth:perspectivesforcontrolleduseofmicroorganismsinagriculture.
ApplMicrobialBiotechnol2009,84:11–18.
14.
MeiC,FlinnBS:Theuseofbeneficialmicrobialendophytesforplantbiomassandstresstoleranceimprovement.
RecentPatentsonBiotechnol2010,4:81–95.
15.
FunkCR,WhiteRH,BreenJP:ImportanceofAcremoniumendophytesinturf-grassbreedingandmanagement.
AgricEcosystEnviron1993,44:215–232.
16.
GhimireSR,CharltonND,CravenKD:Themycorrhizalfungus,sebacinavermifera,enhancesseedgerminationandbiomassproductioninswitchgrass(PanicumvirgatumL).
BioenergyRes2009,2:51–58.
17.
GhimireSR,CharltonND,BellJD,KrishnamurthyYL,CravenKD:Biodiversityoffungalendophytecommunitiesinhabitingswitchgrass(PanicumvirgatumL)growinginthenativetallgrassprairieofnorthernOklahoma.
FungalDiversity2010,47:19–27.
18.
KhanSA,HamayunM,YoonH,KimHY,SuhSK,HwangSK,KimJM,LeeIJ,ChooYS,KongWS,LeeBM,KimJG:PlantgrowthpromotionandPenicilliumcitrinum.
BMCMicrobial2008,8:231–240.
19.
NowakJ,MeiC,LowmanS,ZhaoB,SeilerJ,FlinnB:Developmentofswitchgrass(PanicumvirgatumL.
)formarginallandsbasedongenotypiccompatibilitywithbeneficialbacteria[abstract].
Dalian:InFirstAnnualWorldCongressofBioenergy;2011:25–29.
20.
WelbaumG,SturzAV,DongZ,NowakJ:Managingsoilmicroorganismstoimproveproductivityofagroecosystems.
CritRevPlantSci2004,23:175–193.
21.
CompantS,DuffyB,NowakJ,ClémentC,BarkaEA:Useofplantgrowth-promotingbacteriaforbiocontrolofplantdiseases:principles,mechanismsofactionandfutureprospects.
ApplEnvironMicrobiol2005,71:4951–4959.
22.
CompantS,ReciterB,SessitschA,NowakJ,ClementC,BarkaEA:EndophyticColonizationofVitisvinifereL.
byPlantGrowth-PromotingBacteriumBurkholderiasp.
StrainPsJN.
ApplEnvironMicrobiol2005,71:1685–1693.
23.
GlickBR,PenroseDM,LiJ:Amodelfortheloweringofplantethyleneconcentrationsbyplantgrowthpromotingbacteria.
JTheorBiol1998,190:63–68.
24.
GlickBR:BacterialACCdeaminaseandthealleviationofplantstress.
AdvApplMicrobiol2004,56:291–312.
25.
MattosKA,PáduaVLM,RomeiroA,HallackLF,NevesBC,UlissesTMU,BarrosCF,TodeschiniAR,PreviatoJO,Mendona-PreviatoL:EndophyticcolonizationofricebythedizaotrophicbacteriumBurkholderiakururiensisanditsabilitytoenhanceplantgrowth.
AnAcadBrasCienc2008,80:477–493.
26.
ArshadM,FrankenbergerWT:Microbialproductionofplanthormones.
PlantSoil1991,133:1–8.
Kimetal.
BiotechnologyforBiofuels2012,5:37Page9of10http://www.
biotechnologyforbiofuels.
com/content/5/1/3727.
LazarovitsG,NowakJ:Rhizobacteriaforimprovementofplantgrowthandestablishment.
Hortscience1996,32:188–192.
28.
ConnKL,NowakJ,LazarovitsG:Agnotobioticbioassayforstudyinginteractionsbetweenpotatoesandplantgrowth-promotingrhizobacteria.
CanJMicrobiol1997,43:801–808.
29.
BarkaEA,BelarbiA,HachetC,NowakJ,AudranJC:EnhancementofinvitrogrowthandresistancetograymouldofVitisviniferaco-culturedwithplantgrowth-promotingrhizobacteria.
FEMSMicrobiolLett2000,186:91–95.
30.
NowakJ,AsieduSK,LazarovitsG:Enhancementofinvitrogrowthandtransplantstresstoleranceofpotatoandvegetableplantsco-culturedwithaplantgrowthpromotingrhizobacterium.
InEcophysiologyandPhotosyntheticInVitroCultures.
EditedbyCarreF,ChagvardieffP.
Aix-en-Provence:CEA;2003:173–180.
31.
SessitschA,CoenyeT,SturzAV,VandammeP,BarkaEA,SallesJF,ElsasJD,FaureD,ReiterB,GlickBR,Wang-PruskiG,NowakJ:Burkholderiaphytofirmanssp.
nov.
,anovelplant-associatedbacteriumwithplant-beneficialproperties.
IntJSystEvolMicrobiol2005,55:1187–1192.
32.
WangK,ConnK,LazarovitsG:InvolvementofquinolinatephophoribosyltransferaseinpromotionofpotatogrowthbyaBurkholderiaStrain.
ApplEnvironMicrobiol2006,72:760–768.
33.
WeilharterA,MitterB,ShinMV,ChainPSG,NowakJ,SessitschA:Completegenomesequenceoftheplantgrowth-promotingendophyteBurkholderiaphytofirmansstrainPsJN.
JBacteriol2011,193:3383–3384.
34.
BarkaEA,NowakJ,ClémentS:Enhancementofchillingresistanceofinoculatedgrapevineplantletswithaplantgrowth-promotingrhizobacterium,BurkholderiaphytofirmansstrainPsJN.
ApplEnvironMicrobiol2006,72:7246–7252.
35.
BensalimS,NowakJ,AsieduSK:Aplantgrowthpromotingrhizobacteriumandtemperatureeffectsonperformanceof18clonesofpotato.
AmJPotatoRes1998,75:145–152.
36.
FrommelMI,NowakJ,LazarovitsG:Growthenhancementanddevelopmentalmodificationofinvitrogrownpotato(Solanumtuberosumspp.
tuberosum)asaffectedbyanonfluorescentPseudomonassp.
PlantPhysiol1991,96:928–936.
37.
SharmaV,NowakJ:Enhancementofverticilliumwiltresistanceintomatotransplantsbyinvitroco-cultureofseedlingswithaplantgrowthpromotingrhizobacterium(Pseudomonassp.
strainPsJN).
CanJMicrobiol1998,44:528–536.
38.
BarkaEA,GogniesS,NowakJ,AudranJ,BelarbiA:InhibitoryeffectofendophytebacteriaonBotrytiscinereaanditsinfluencetopromotegrapevinegrowth.
BiolControl2002,24:135–142.
39.
ReiterB,PfeiferU,SchwabH,SessitschA:ResponseofendophyticbacterialcommunitiesinpotatoplantstoinfectionwithErwiniacaratovorasubsp.
atroseptica.
ApplEnvironMicrobiol2002,68:2261–2268.
40.
WhippsJM:Microbialinteractionsandbiocontrolintherhizosphere.
JExpBot2001,52:487–511.
41.
LongHH,SchmidtDD,BaldwinIT:Nativebacterialendophytespromotehostgrowthinaspecies-specificmanner;phytohormonemanipulationsdonotresultincommongrowthresponse.
PLoSOne2008,3:e2702.
42.
PenroseDM,GlickBR:MethodsforisolatingandcharacterizingACCdeaminase-containingplantgrowthpromotingrhizobacteria.
PhysiolPlant2003,118:10–15.
43.
NowakJ,AsieduSK,BensalimS,RichardsJ,StewartA,SmithC,StevensD,SturzAV:Fromlaboratorytoapplications:challengesandprogresswithinvitrodualculturesofpotatoandbeneficialbacteria.
PlantCellTissOrgCult1998,52:97–103.
44.
NowakJ,VeilleuxRE,NowakJ,TurgeonS:Primingfortransplantstressresistanceininvitropropagationviaplantletbacterization.
ActaHort2007,748:65–75.
45.
PillayVK,NowakJ:Inoculumdensity,temperature,andgenotypeeffectsoninvitrogrowthpromotionandepiphyticandendophyticcolonizationoftomato(LycopersiconesculentumL.
)seedlingsinoculatedwithapseudomonadbacterium.
CanJMicrobiol1997,43:354–361.
46.
MurashigeT,SkoogF:Arevisedmediumforrapidgrowthandbioassayswithtobaccotissuecultures.
PhysiolPlant1962,15:473–497.
doi:10.
1186/1754-6834-5-37Citethisarticleas:Kimetal.
:Growthpromotionandcolonizationofswitchgrass(Panicumvirgatum)cv.
AlamobybacterialendophyteBurkholderiaphytofirmansstrainPsJN.
BiotechnologyforBiofuels20125:37.
SubmityournextmanuscripttoBioMedCentralandtakefulladvantageof:ConvenientonlinesubmissionThoroughpeerreviewNospaceconstraintsorcolorgurechargesImmediatepublicationonacceptanceInclusioninPubMed,CAS,ScopusandGoogleScholarResearchwhichisfreelyavailableforredistributionSubmityourmanuscriptatwww.
biomedcentral.
com/submitKimetal.
BiotechnologyforBiofuels2012,5:37Page10of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37
AlamobybacterialendophyteBurkholderiaphytofirmansstrainPsJNSeonhwaKim1,ScottLowman1,2,GuichuanHou4,JerzyNowak2,BarryFlinn1,2,3andChuanshengMei1,2,3*AbstractBackground:SwitchgrassisoneofthemostpromisingbioenergycropcandidatesfortheUS.
Itgivesrelativelyhighbiomassyieldandcangrowonmarginallands.
However,itsyieldsvaryfromyeartoyearandfromlocationtolocation.
Thusitisimperativetodevelopalowinputandsustainableswitchgrassfeedstockproductionsystem.
Oneofthemostfeasiblewaystoincreasebiomassyieldsistoharnessbenefitsofmicrobialendophytes.
Results:Wedemonstratethatoneofthemoststudiedplantgrowthpromotingbacterialendophytes,BurkholderiaphytofirmansstrainPsJN,isabletocolonizeandsignificantlypromotegrowthofswitchgrasscv.
Alamounderinvitro,growthchamber,andgreenhouseconditions.
Inseveralinvitroexperiments,theaveragefreshweightofPsJN-inoculatedplantswasapproximately50%higherthannon-inoculatedplants.
Whenone-month-oldseedlingsweregrowninagrowthchamberfor30days,thePsJN-inoculatedAlamoplantshadsignificantlyhighershootandrootbiomasscomparedtocontrols.
Biomassyield(dryweight)averagedfromfiveexperimentswas54.
1%higherintheinoculatedtreatmentcomparedtonon-inoculatedcontrol.
Similarresultswereobtainedingreenhouseexperimentswithtransplantsgrownin4-gallonpotsfortwomonths.
Theinoculatedplantsexhibitedmoreearlytillersandpersistentgrowthvigorwith48.
6%higherbiomassthancontrols.
WealsofoundthatPsJNcouldsignificantlypromotegrowthofswitchgrasscv.
Alamoundersub-optimalconditions.
However,PsJN-mediatedgrowthpromotioninswitchgrassisgenotypespecific.
Conclusions:OurresultsshowB.
phytofirmansstrainPsJNsignificantlypromotesgrowthofswitchgrasscv.
Alamounderdifferentconditions,especiallyintheearlygrowthstagesleadingtoenhancedproductionoftillers.
Thisphenomenonmaybenefitswitchgrassestablishmentinthefirstyear.
Moreover,PsJNsignificantlystimulatedgrowthofswitchgrasscv.
Alamoundersub-optimalconditions,indicatingthattheuseofthebeneficialbacterialendophytesmayboostswitchgrassgrowthonmarginallandsandsignificantlycontributetothedevelopmentofalowinputandsustainablefeedstockproductionsystem.
Keywords:Bacterialendophyte,BurkholderiaphytofirmansstrainPsJN,Colonization,Growthpromotion,Biomassincrease,Switchgrasscv.
Alamo*Correspondence:chuansheng.
mei@ialr.
org1InstituteforSustainableandRenewableResource,InstituteforAdvancedLearningandResearch,150SlaytonAve,Danville,VA24540,USA2DepartmentofHorticulture,VirginiaPolytechnicInstituteandStateUniversity,Blacksburg,VA24601,USAFulllistofauthorinformationisavailableattheendofthearticle2012Kimetal.
;licenseeBioMedCentralLtd.
ThisisanOpenAccessarticledistributedunderthetermsoftheCreativeCommonsAttributionLicense(http://creativecommons.
org/licenses/by/2.
0),whichpermitsunrestricteduse,distribution,andreproductioninanymedium,providedtheoriginalworkisproperlycited.
Kimetal.
BiotechnologyforBiofuels2012,5:37http://www.
biotechnologyforbiofuels.
com/content/5/1/37BackgroundIncreasingconcernoverforeignenergysupplies,globalgreenhousegasemissionsandtheneedforruraleco-nomicdevelopmenthasdriventheinterestinsustain-ablebiomassproductionforbioenergyandbio-products.
Ithasbeensuggestedthatby2025,theworldenergyde-mandwilllikelybeincreasedbymorethan50%[1,2].
Thisdemand,andsocietalconcernsabouttheenviron-mentalimpactofburningfossilfuelsarekeyfactorsstimulatingthedevelopmentofnationalandregionalstrategiesaimedatthegrowthofrenewableenergysup-plies,primarilyfocusedonbiofuels.
Toreducethereli-anceonfossilfuels,theUSA,theworld'smajorenergyconsumer,releasedtheEnergyIndependenceandSecur-ityActof2007thataimstoincreasetheproductionofrenewablefuelsfrom9.
0billiongallonsin2008to36billiongallonsby2022[3].
TherecentUSDA/DOENa-tionalBiofuelsActionPlan[4]hashelpedtodelineatethepriorityareasrequiredtoacceleratesustainablebio-fuelindustrydevelopment.
Withinthisdocument,Ac-tionArea2wasidentifiedasfeedstockproductionandimprovement.
Variousfeedstocks,suchasperennialrhi-zomatousgrasses,canprovidesourcesoflignocellulosicbiomass,servingasnewsourcesofcropgrowthandin-comeforregionalfarmers.
Oneofthemostpromisingfeedstockscapableofcon-tributingtotherealizationofUSrenewableenergygoalsisthecommonperennialgrass,switchgrass(PanicumvirgatumL.
)[5].
Thisnativeprairiegrass,consistingofadiversegermplasm[6],cangrowonmarginallandsunderlowinputsofwaterandagrochemicals[7],sothatitscultivationdoesnotcompetewithfoodcropsforlandandotherresources.
Duetoitslargerootsystemandfaststandregrowth,switchgrasshasotherpositiveenvir-onmentaleffects,includingthepreventionofsurfacerunoffandsoilerosion,carbonsequestration,andtheprovisionofawildlifehabitat[5,8].
Switchgrassculti-vatedlandsalsohadmuchhighertotalsoilorganiccar-bondepositsthanlandscultivatedwithannualcrops,suchascornandwheat[9,10].
Theeconomicsofbiofuelproductionishighlydependentonfeedstockcostandconversiontechnology[1,6].
Thedevelopmentofimprovedswitchgrassvarietiesforlow-costproductiononmarginallandsisoneprerequisiteforthesuccessofthebioenergyprogram[5,11].
Onesuchap-proachinvolvestheuseofbeneficialmicroorganisms,suchasendophytes,whichformintimateassociationswithplants[12,13].
Endophytes,bothfungalandbacterial,havebeentargetedasmechanismstoenhanceplantcharacteris-ticsforcommercialuses[14].
Thecolonizationofgrassesbyfungalendophytesforperformanceenhancementiswelldocumented[15],includingtheirusewithswitchgrass[16,17].
However,toourknowledge,onlyonestudyhasreportedgrowthpromotionofabioenergyfeedstockgrass(Miscanthusxgiganteous)seedlingsbyabacterialendophyte(Herbaspirillumfrisingense)[18].
Akeycom-ponentofourbioenergycropresearchprograminvolvestheutilizationofbeneficialbacterialendophytesthatformstableandpersistentassociationswithswitchgrass,asthemechanismtoimprovebiomassyieldandenhancestresstoleranceunderlow-inputproductionsystems[19].
Bene-ficialbacterialendophytesarenaturallyoccurringsoilmicroorganismsthatcanpenetrateplantrootsandtrans-locatetotheabovegroundorgansand,uponcolonization,affectplantgrowth,health,andproductivity[12,20-22].
Althoughthemolecularmechanismsofbeneficialendophyte-hostplantinteractionsarelargelyunknown,severalstudieshavedemonstratedthatendophytescanpromoteplantgrowthbyenhancingtheplant'scapacityfornutrientacquisition,betterwatermanagement,and/orresistancetoabioticandbioticstressesviaregulationofhormones[12-14,20,21].
Forinstance,1-aminocyclo-propane-1-carboxylicacid(ACC)deaminaseproducedbyendophyteslowerstheethylenelevelsinhostplants,redu-cingtheirresponsetoabioticandbioticstress,andbychangingrootmorphology,leadingtostimulationofplantgrowth[13,23,24].
Manyknownendophytesalsopromoteplantgrowthbyproducinggibberellicacid(GA3),indole-3-aceticacid(IAA)[18,25],orcytokinins[26,27].
BurkholderiaphytofirmansstrainPsJNhasbeenfoundtobeahighlyeffectiveplantgrowthpromotingbacterialendophyte,withabroadhostrangeincludingpotatoes,tomatoes,andgrapevines[21,27-32].
Inaddition,itsgenomehasrecentlybeensequenced[33],providingthegenomicresourcesneededtodevelopanunderstandingofthemechanismsassociatedwiththisendophyte'sabilitytopromoteplantgrowth.
PsJNpro-ducesahighlevelofACCdeaminase[31],enhanceshostplantcold[34]andheat[35]stresstolerance,improveswatermanagement[36]andplantresistancetopathogens[37,38].
Inthisstudy,wereportgrowthpromotionofswitchgrasscv.
AlamobyBurkholderiaphytofirmansstrainPsJNunderinvitro,growthcham-ber,andgreenhouseconditions.
Toourknowledge,thisisthefirstreportdetailingtheswitchgrass-PsJNinteraction.
ResultsPsJNendophyticassociationwithswitchgrassAlamoTheendophyticcolonizationofswitchgrassbyBurkhol-deriaphytofirmansstrainPsJN-GFPwasvisualizedusingconfocalmicroscopy(Figure1).
Undertheappropriateillumination,thePsJN-GFPcouldbeclearlyobservedin-sidetherootsofPsJN-inoculatedplants3daysafterin-oculation,whilenofluorescencewasobservedinrootsofbuffer-inoculatedcontrolplants.
ThetiterofPsJN-GFPininoculatedplantswasalsodeterminedusingtis-suehomogenatesfromvarioustissues(root,leafandKimetal.
BiotechnologyforBiofuels2012,5:37Page2of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37sheath)atdifferenttimes(Table1).
Theendophyteini-tiallyinfectedandcolonizedplantroots,andby7dayspost-inoculation,PsJNtiterswerestillhighestintheroot.
However,thetitersincreasedsignificantlyinothertissuesbyday14,indicatingtranslocationtoleavesandsheaths.
EffectsofPsJNonAlamogrowthinvitroYoungswitchgrassseedlingswerepreparedandinocu-latedasdescribedinMaterialsandMethods,andthenon-inoculatedandinoculatedplantswereanalyzedaftergrowthinvitroforonemonth.
TheresultshowedthatPsJNsignificantlyandrepeatedlypromotedAlamorootandshootgrowth,witha35.
6%increaseinshootlength,a32.
8%increaseinrootlength,andan83.
6%in-creaseinfreshweight,comparedtothenon-inoculatedplants(Figure2).
Afterseveralreplications,theaveragefreshweightofthePsJN-inoculatedplantswasalwaysapproximately50%higherthannon-inoculatedplants.
EffectsofPsJNonAlamogrowthinagrowthchamberenvironmentAsdescribedabove,PsJNsignificantlyenhancedswitch-grasscv.
Alamogrowthinvitro.
Wenextassessedtheim-pactofPsJNongrowthundersoilconditions.
One-month-oldinvitrogrownAlamo(controlandPsJN-inoculatedseedlings)weretransferredtoaflatwith72cavitiesfilledwithsoilandgrowninagrowthchamberunder28/22°Cday/nighttemperatureswith16hlightperiodforonemonth.
ThePsJN-inoculatedplantsshowedsignificantgrowthincreasescomparedtocontrolplantsinshootlengthandfresh/dryweights(Figure3).
Thegrowthcham-berexperimentswererepeated5times,andtheaveragedatafrom5experimentsshowedsignificantgrowthpro-motionbyPsJN,witha46.
3%,anda54.
1%increaseinfreshweightanddryweight,respectively.
Thetotaldryweightincrease(54.
1%)byPsJNwasmorethanthetotalfreshweightincrease(46.
3%),indicatingthatthePsJN-inoculatedplantsaccumulatedmorebiomass.
EffectsofPsJNonAlamogrowthinthegreenhouseNon-inoculatedandPsJN-inoculatedplantswerealsogrownundergreenhouseconditionstodeterminegrowthenhancementpersistence.
TheplantsinoculatedwithPsJNandgrowninvitrofor25daysweretransferredto4-gallonpotswith5plantsineachpotandgrowninthegreen-house.
TheplantsinoculatedwithPsJNexhibitedsustainedgrowthvigor,astheyweresignificantlytaller,andmoretil-lersdevelopedearlycomparedwiththenon-inoculatedcontrolplants(Figure4).
Followingonemonthofgrowthinthegreenhouse,thePsJN-inoculatedplantshad76.
2%moretillersthanthecontrolplants.
TheplantswereFigure1Confocalimagesofrootstaken3daysfollowingswitchgrasscv.
AlamoinoculationwithPsJN-GFP,showingbacterialcolonizationinsidetheroots.
(A):Controland(B):PsJN-inoculatedplants.
(C):Controland(D):PsJN-inoculatedplants.
Ineachsection(a,b,c,d),leftpanelswereobservedunderfluorescentlightandrightpanelsundervisiblelight.
Thebarsrepresent100μm(AandB)and20μm(CandD).
Table1Colony-formingunits(CFU)ofroot,leaf,andsheathtissuesatdifferenttimesafterPsJN-GFPinoculationDaysafterPsJN-PlanttissuesAverageCFU/gGFPinoculationFreshweight3All(Roots,leaves,andsheath)4.
2X1057Roots7.
6X105Leaves2.
6X10314Roots3X104Sheaths1.
3X105Leaves1.
2X105Kimetal.
BiotechnologyforBiofuels2012,5:37Page3of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37harvestedfollowinggrowthfortwomonths,andthebio-massyielddetermined(Figure5).
ThePsJN-inoculatedplantswererepeatedlysignificantlyhigherinbiomassyield,witha36.
8%increaseinfreshweightanda57.
1%increaseindryweight.
EffectsofPsJNonAlamogrowthinsub-optimalconditionsInordertodevelopalowinputandsustainableswitch-grassfeedstockproductionsystemutilizingthebeneficialbacterialendophyte,wetestedgrowthperformanceofPsJN-inoculatedplantwithunfertilizedfieldsoil,inaglasshouseunderambientconditionsduringtheFall,whenthetemperaturewasnotoptimalforswitchgrassgrowth.
TheresultsshowedthatPsJN-inoculatedplantsproducedtwicethetotalbiomassofcontrols(Figure6).
DirectinoculationofswitchgrassseedswithPsJNInordertoexploreapracticalwaytoinoculateswitch-grasswiththebacterialendophyte,westerilizedswitch-grassseedsasdescribedinMaterialsandMethods,placedthesterilizedseedsonwetfilterpaperfor3-5daysinanincubatorat25°C,andtheninoculatedthegerminatingseedswithdifferentconcentrationsofendophyteinocu-lumtodeterminetheoptimalinoculationconcentration(OD600at0.
1-0.
5).
TheplantsinoculatedwithPsJNatOD600of0.
1,0.
25,and0.
5exhibited28.
7%,55.
0%and80.
1%increasesindryweight,respectively,comparedtonon-inoculatedplantsaftergrowninvitrofor25daysandingrowthchamberforanothermonth.
APsJNconcentra-tionof0.
5wasthemosteffectiveatpromotingbiomassincrease(Figure7),andnobiomassdifferencewasobservedbetweenthe0.
1treatmentandcontrol.
Endophyteinfectionandcolonizationofseedsaredependentonendophyteconcentrationandthestatusofseedimbibitions.
So,inordertofacilitateinfectionandcolonizationbythebacterialendophyte,thesterilizedseedswereimbibedinwaterfor1,2,3,or4days,andtheninoculatedwithPsJNatanOD600of0.
5or1.
0,sinceanOD600of0.
5wasthemosteffectiveasdescribedabove.
ThePsJN-inoculatedseedswereplacedinanin-cubatorat25°Cwitha16hlightperiodfor25days,andtransferredtosoilandgrowninagrowthchamberfor37days.
Theresultsindicatedthatplantsfromtheseedsimbibedfor2daysandtheninoculatedwithanOD600of0.
5hadthehighestdryweight,witha55%increasecomparedtonon-inoculatedcontrolplants(Figure8).
GenotypicresponsestoPsJNAsdescribedabove,PsJNwasabletostimulategrowthinswitchgrasscv.
Alamo.
ToassesstheinfluenceofFigure2EffectsofendophytePsJNinoculationonswitchgrasscv.
Alamogrowthinvitro.
Datawereobtainedafterplantsweregrowninincubatorfor36days.
Samplenumberwas25,and**meanssignificantdifferenceat0.
01levelbetweenPsJNandcontrolusingstudentT-test.
Figure3EffectsofendophytePsJNinoculationonswitchgrasscv.
Alamogrowthingrowthchamber.
TheseedlingswereinoculatedwithPsJNandgrowninvitroforonemonth,thentransferredtosoilandgrowningrowthchamberforanothermonth.
Dryweightwasdeterminedaftersamplesweredriedinovenat65°Cforoneday.
Samplenumberwas36,and**meanssignificantdifferenceat0.
01levelbetweenPsJNandcontrolusingstudentT-test.
Kimetal.
BiotechnologyforBiofuels2012,5:37Page4of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37plantgenotypeonthisresponse,sevenotherswitchgrasscultivarsweretestedfortheirgrowthresponsestoPsJN.
AsshowninTable2,growthpromotionbyPsJNwasgenotype-dependent.
Theswitchgrasscvs.
Forestburg,Nebraska,andBlackwellwereallresponsivetoPsJN,withmeasuredsignificantgrowthincreasesof60.
1%,26.
8%,and23.
0%,respectively,whilethecvs.
Cave-in-Rock,Sunburst,Shelton,andShawneedidnotexhibitgrowthpromotioninresponsetoPsJNundersimilarconditions.
PreliminaryresultfromCave-in-Rockbio-assayindicatedthatPsJNtiterswerenotsustainedafterinoculationandweremuchlowerinthenon-responsiveplantsfollowinginoculation.
DiscussionInthepresentstudy,wedemonstratedtheabilityofB.
phytofirmansPsJNtocolonizeandpromotegrowthinswitchgrasscv.
Alamo.
ThreedaysfollowingPsJNinocu-lation,wecouldclearlyvisualizebacterialcolonizationinsidetherootsunderconfocalmicroscopy.
Thebacter-ialpopulationinsidetherootswasinitiallymuchhigherthanthatoftheleavesandsheaths,andthebacterialendophytewassubsequentlytranslocatedverticallytotheupperleavesthroughtheleafsheath.
Theseresultsweresimilartothatreportedforgrapevine[22]andpo-tato[39],wherePsJNwastransportedthroughtheinter-iorvascularsystem,fromrootxylemvesselstotheupperpartsoftheplants.
Thisisacriticalfirststepintheendophyticbacteria-plantinteraction[40].
Weobservedsignificantgrowthpromotionofcv.
AlamobyPsJN,underbothinvitroandsoilconditions.
Ourstudyshowedtotalfreshweightandtotaldryweightoftheinoculatedplantswasincreasedby45%and55%respect-ivelycomparedwiththenon-inoculatedcontrolplantswhentheinoculatedseedlingsweregrowninvitroandthentransferredtosoilandgrowningrowthchamberforonemonth.
Similarresultshavebeenobtainedin4-gallonpotsunderourgreenhouseconditions.
OtherstudieshavereportedlevelsofgrowthpromotionbyPsJN,withgrapevineshowinga6-foldincreaseintotalbiomass[34],andpotatoshowinganapproximate2-foldincreaseinrootandhaulmbiomass[36]overcontrols.
ThemechanismofplantgrowthbyB.
phytofirmansPsJNhasbeenreported[21]andattributedtotheabilityFigure4PlantgrowthandtillerdevelopmentaftercontrolandPsJNinoculatedplantsweretransferredto4-gallonpotsandgrowningreenhouse.
Figure5GrowthpromotionpersistenceofAlamoingreenhousebyPsJNinoculation.
TheseedlingswereinoculatedwithPsJNandgrowninvitroforonemonth,thentransferredto4-gallonpotwith5plants/potandgrowningreenhousefortwomonths.
Dryweightwasdeterminedaftersamplesweredriedinovenat65°Cforoneday.
Samplenumberwas25,and**meanssignificantdifferenceat0.
01levelbetweenPsJNandcontrolusingstudentT-test.
Kimetal.
BiotechnologyforBiofuels2012,5:37Page5of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37ofPsJNtoproducehighlevelsofACCdeaminaseactiv-ity,whichdegradesACCtoammoniaandα-ketobutyrate[41],whichisacommoncharacteristicofplantgrowthpromotingbacteria.
ACCistheprecursortoethylene,aplantstresshormone;hence,thereducedethylenelevelinPsJN-colonizedplantswillpromoteplantgrowth.
AccordingtothereportbyPenroseandGlick(2003)[42],ACCactivityover20nmolα-ketobutyrate/h/mgissufficienttopromotehostplantgrowth,andPsJNhasbeenshowntocontain308nmolα-ketobutyrate/h/mgofACCdeaminaseactivity[31].
Al-thoughseveralstudieshavereportedtheinteractionbe-tweenthisendophyteandhostplantsforgrowthpromotion,moststudieshavereportedinvitrodata[22,27,28,36].
Ourresultswithunfertilizedfieldsoil,inaglasshouseunderambientconditionsduringtheFall,whenthetemperaturewasnotoptimalforswitchgrassgrowth(Figure6)impliedthepotentialbenefitofswitch-grassinoculatedwithPsJNforgrowthonmarginallandsandsub-optimalgrowthconditions.
Althoughourresultsindicatepotentialbenefitofbacterialendophytesinswitchgrassunderinvitro,growthchamberandgreen-houseconditions,muchworkstillneedstobedoneinassessinggrowthpromotionofswitchgrassbyendo-phytesinthefieldsincethereismuchmorecompetitionbetweenendophytesandothermicroorganismsinadditiontomanyotherunfavorableconditions.
Whileourinitialstudieswerecarriedoutwithcv.
Alamo,wetestedtheutilityofPsJNasagrowth-promotingendo-phytewithotherswitchgrasscultivars.
Ourresultsindi-catedthatspecificgenotypeeffectsexisted,withsomegenotypesbeinghighlyresponsivetothegrowthpromo-tiveeffectsofPsJN,andothersnot.
Similargenotypeeffectshavebeenreportedbyothers.
ItwasreportedthatthepotatoresponsetoPsJNinvolvessomeformofgen-eticcontrol,assomepotatocultivarsdisplaythebenefi-cialresponsetotheendophyte,whileothersdonot[35,43,44].
Thetypicalinvitrophenotypeforastronglyresponsivecultivarwascharacterizedbyamassive,well-branchedrootsystemandafterthefirst3-4weeksinFigure6GrowthpromotionofAlamobyPsJNinoculationinsub-optimalconditions.
TheseedlingswereinoculatedwithPsJNandgrowninvitroforonemonth,thentransferredto4-gallonpotswith5plants/potwithunfertilizedfieldsoilandgrownintheglasshouseunderambientconditionsfor2.
5monthsinthelateFallof2010.
Dryweightwasdeterminedaftersamplesweredriedinovenat65°Cforoneday.
Samplenumberwas25,and**meanssignificantdifferenceat0.
01levelbetweenPsJNandcontrolusingstudentT-test.
Figure7EffectsofdifferentconcentrationsofPsJNonswtchgrasscv.
Alamogrowthwithdirectseedinoculation.
Thesurface-sterilizedseedswereinfectedwithdifferentconcentrationsofPsJNandgrowninvitrofor17days,thentransferredto72-cavitytraysandgrowninagrowthchamberfor50days.
*and**meansignificantdifferenceat0.
05and0.
01levelsrespectivelybetweenPsJNandcontrolusingstudentT-test.
Kimetal.
BiotechnologyforBiofuels2012,5:37Page6of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37culture,theplantletwasdevelopmentallymoreadvancedthanthenon-bacterizedcontrols.
Stemsweresturdier,withmorelignindepositsaroundthevascularsystem,andtheplantletsdevelopedmoreroothairsandmoreandlargerleaftrichomes[43].
WealsonoticedPsJN-inoculatedswitchgrassplantsweredevelopmentallyadvanced(unpublisheddata).
Suchenhancementswerenotapparentforthepoorly-responsivecultivars.
WealsoobservedsomeofthesephenotypicdifferencesbetweenPsJN-responsiveandnon-responsiveswitchgrassculti-vars.
AdditionalworkillustratingthegeneticcontrolofthebeneficialresponsetotheendophyteusedmonoploidpotatolinesderivedfromanthercultureofanadapteddiploidSolanumphurejaclone,BARD1-3[44].
Thedip-loidantherdonor,BARD1-3,exhibitedabacterizationresponsecomparabletoRedPontiac,whilemonoploidlinesexhibitedaresponsetoPsJNrangingfromfavorabletounfavorabletoneutral.
Theassumptionherewasthattheresponserangeofthemonoploidpopulationwasduetothesegregationofallelesforgenesinvolvedinregulat-ingthepositiveornegativeinteractionwithPsJN.
Thepotato/PsJNstudieshavebeenthemostcharacter-ized,andcarriedoutwithmaterialclonallypropagatedvianodalsections,inwhichasingleinoculationissuffi-cienttoinitiatecolonizationthroughthexylemtissue,Figure8EffectsofdifferentconcentrationsofPsJNonswitchgrasscv.
Alamogrowthwithdirectinoculationofseedsthatwereimbibedinwaterfordifferentdays(1-4days).
TheseedswereinfectedwithdifferentconcentrationsofPsJNandgrowninvitrofor25days,thentransferredto72-cavitytraysandgrowninagrowthchamberfor37days.
Samplenumberwas72foreachtreatment.
*and**meansignificantdifferenceat0.
05and0.
01levelsrespectivelybetweenPsJNandcontrolusingstudentT-test.
Table2EffectsofB.
phytofirmansPsJNonplantgrowthindifferentswitchgrasscultivarsCultivarsTreatmentNo.
plantsRootlength(cm)Shootlength(cm)Totalfreshweight(mg)PsJN/control(%)aShawneeControl242.
38.
450.
1PsJN242.
17.
858.
8117.
4p-valueb0.
20370.
20490.
0717NebraskaControl241.
510.
340.
9PsJN241.
511.
651.
9126.
8p-value0.
41600.
06560.
0055ForestburgControl341.
68.
633.
7PsJN301.
812.
754.
0160.
1p-value0.
19860.
00000020.
0000001SheltonControl283.
314.
1117.
7PsJN283.
012.
0135.
3115.
0p-value0.
15790.
00670.
0907BlackwellControl281.
49.
952.
6PsJN281.
511.
364.
7123.
0p-value0.
29700.
09980.
0543SunburstControl300.
88.
533.
0PsJN280.
910.
426.
480.
0p-value0.
29850.
01370.
0731Cave-in-RockControl332.
814.
7107.
8PsJN343.
113.
8113.
4105.
2p-value0.
23170.
40520.
5613aPsJN/control(%)meanstotalfreshweightofPsJNinoculatedplantsoverthatofcontrolplants.
bp-valuemeansdegreesofstatisticalsignificancebetweenPsJNtreatmentandcontrolusingstudentT-test.
Kimetal.
BiotechnologyforBiofuels2012,5:37Page7of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37eventuallyspreadingtotheupperleaves[36].
Bacteriallevelsmustreachathresholdpopulationwithintheplantbeforetheyareeffective[45]withadirectrelation-shipbetweenplantletgrowthenhancementandPsJNcolonizationofbothinteriorandexteriorsurfaces[44].
ThePsJNcolonizationprofilesforaresponsiveandpoorlyresponsivecultivarover8tissueculturegenera-tionsrevealedbacterialloadsoneorderofmagnitudegreaterforshoot/rootsurfaceandinteriorcolonizationintheresponsivecomparedtothepoorlyresponsivecul-tivar.
Furthermore,theresponsivecultivarexhibitedincreasedcolonizationoversuccessivegenerations,whilethepoorlyresponsivecultivarexhibiteddecliningbacter-ialpopulationsoversuccessivegenerations.
Wearecur-rentlyassessingthelevelofcolonizationinswitchgrasscultivarsresponsiveandnon-responsivetoPsJNtode-terminethedegreeofsimilaritybetweenswitchgrassandpotatoresponsestotheendophyte.
Atpresent,themechanismsgoverningB.
phytofirmansPsJNgenotype-specificityingrowthpromotionofswitchgrass(andotherplants)areunknown,althoughwearecurrentlyusinglargescalegeneexpressionanalysestodeterminethedifferencesintheswitchgrassmolecularresponsesbetweendifferently-respondingcultivars.
Insummary,theresultsreportedhereillustratetheabilityofB.
phyto-firmansPsJNtoinfectandcolonizeresponsiveswitch-grass(Panicumvirgatum)cultivarslikeAlamo,andtopromoteplantgrowth.
Thisstudylaysthefoundationtodevelopalowinputandsustainableswitchgrassfeed-stockproductionsystemonmarginallandsusingthis,andother,beneficialbacterialendophytes.
ConclusionsResultsobtainedwithgrowthpromotionofswitchgrasscv.
AlamobyB.
phytofirmansPsJNundervariousgrowthconditions,sub-optimalinparticular,indicatethepotentialforutilizationofbeneficialbacterialendo-phyteinswitchgrassestablishmentinthefirstyearandinthedevelopmentoflowinputandsustainableswitch-grassfeedstockproductionsystem.
Inthefuture,themechanismsofgrowthpromotionneedtobeelucidatedwithmolecularbiologyandfunctionalgenomicstode-veloptoolsformolecularbreedingforbeneficialplant-microbialassociations.
MethodsPlantmaterialsSwitchgrass(PanicumvirgatumL.
)cvs.
AlamoandCave-in-RockseedswerepurchasedfromWarnerBroth-ersSeedCo.
(Lawton,OK),andotherswitchgrassseedswerekindlyprovidedbyDr.
BingyuZhao(DepartmentofHorticulture-VirginiaTech,Blacksburg,VA).
Switch-grassseedsweresurface-sterilizedbytreatmentwith70%ethanolfor2min,rinsed3Xwithdistilledwater,de-huskedfor30minwith60%H2SO4withstirring,washed3Xwithdistilledwater,sterilizedwith0.
4Mso-diumhypochlorite(50%commercialbleachsolutioncontaining6%sodiumhypochlorite)containing0.
1%Triton100for30minfollowedby5Xrinsewithsterile,deionized,distilledwater(ddH2O).
BacterialendophyteandcultureconditionsBurkholderiaphytofirmansstrainPsJN[31]anditsPsJN-GFPderivative[22]wereobtainedfromDr.
AngelaSessitsch(AustrianInstituteofTechnology,Seibersdorf,Austria).
ThecultureswerestreakedonKing'sB(KB)solidmediumasdescribedin[45].
Inoculumwaspro-ducedbytransferringoneloopofPsJNfrom2-day-oldculturesto5mlKBbrothina15-mlculturetube,fol-lowedbyincubationat28°Conashaker(150rpm)over-night.
FivemloftheovernightPsJNculturewasaddedto45mlKBbrothina250-mlErlenmeyerflaskandgrownto0.
7OD600.
Bacterialcellswerethencollectedbycentrifugationat3,500rpmfor7minat4°C,andre-suspendedinPBSbuffer(10mMNaH2PO4containing0.
8%NaC1,pH6.
5)afterwhichtheOD600wasadjustedwithPBSbufferto0.
5,unlessdescribedotherwise.
SeedlinginoculationwithPsJNandplantgrowthresponsesSurface-sterilizedseedsweregerminatedinpetri-dishesfor7daysat25°C,underwhitefluorescentlight(67μmolm-2s-1),16hphotoperiod,onaswitchgrassgrowthmediumconsistingofMSsaltsandvitamins[46],30g/lmaltoseand3g/lphytogel,pH5.
8.
TheroottipsoftheyoungseedlingswerecutpriortoPsJNinocu-lationtofacilitatebacterialpenetration[45].
Forthedir-ectseedinoculationsurface-sterilizedseedswereplacedonwetfilterpaperfor3-5daysinanincubatorat25°Cwith16hphotoperiod(whitefluorescentbulbsat67μmolm-2s-1)followedbysoakinginPsJNsolution(0.
5ofOD600)for1min.
Controlseedlings/seedsweretreatedwithPBSbufferalone.
Thetreatedseedling/seedswereblot-driedwithsterilepapertowel,placedonswitchgrassgrowthmediuminGA7Magentavessels(Sigma-Aldrich)containing50mlofmediaand5seed-lingsorgerminatingseedspervessel,andgrownforonemonthintheincubatorasabove.
Rootandshootlength,andseedlingfreshweightwerethendetermined,andtheplantstransferredtoasoilmixcomposedof2/3Miracle-GroWPottingMix(ScottsMiracle-GroCom-pany,Marysville,Ohio)and1/3Arabidopsisgrowingmedia(LehleSeeds,RoundRock,Texas).
Plantsweregrownin72-cavitytraysinagrowthchamberata28/22°Cday/nighttemperature,16hphotoperiod(whitefluorescentbulbsat88μmolm-2s-1)for30days,or4-gallonpotsinthegreenhouse.
Kimetal.
BiotechnologyforBiofuels2012,5:37Page8of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37PsJNcolonizationTheplantsinoculatedwithPsJN-GFPwereexaminedunderafluorescentstereomicroscope(ModelSZX-ILLD2-100;Olympus,Tokyo,Japan)equippedwithaGFPfilter(BP460-490,Olympus,Tokyo,Japan)andtheZeiss510laserscanningconfocalmicroscope(LSCM)(CarlZeiss,Inc.
,Thornwood,NY)toobservecolonization.
Forbioassays,thecontrolandPsJN-GFPinoculatedplantsweresurface-sterilizedwith0.
032Msodiumhypo-chloritefor1min,thenwashed4Xwithsteriledistilledwater.
FiftyμlofthefinalwashwasplatedonsolidKBmediumtoconfirmeffectivenessofsurfacesterilization.
Root,leafandsheathpartswerethenseparated,eachweighed,andgroundwithmortarandpestlein1mlster-iledistilledwater.
Thehomogenateswerethencentrifugedat2000rpmfor3min,andthesupernatantdilutedto1:10,1:100,and1:1000withdistilledwater.
Fiftyμlsam-plesoftheseriallydilutedsolutionswerespreadonsolidKBmedium.
Theplateswereincubatedfor3daysat28°CinthedarkandthenumberofGFP-positivecoloniesdeterminedusingfluorescencestereomicroscopyasdescribedabove.
CompetinginterestsTheauthorsdeclarethattheyhavenocompetinginterests.
Authors'contributionsSKandSLconductedexperimentsanddraftedthemanuscript.
GHcarriedoutconfocalimaginganddataanalysisandreviewedthemanuscript.
CM,JNandBFdesignedexperimentsandwrotethepaperwithinputfromallauthors.
Allauthorsreadandapprovedthefinalmanuscript.
AcknowledgementsThisworkwasfundedthroughSpecialGrants(2003–38891–02112,2008-38891-19353and2009-38891-20092)andHATCHfunds(ProjectNo.
VA-135816)fromtheUnitedStatesDepartmentofAgriculture,theOfficeofScience(BER),U.
S.
DepartmentofEnergyforPlantFeedstockGenomicsforBioenergyProgram(DE-SC0004951),andoperatingfundsfromtheCommonwealthofVirginiatotheInstituteforAdvancedLearningandResearch.
Authordetails1InstituteforSustainableandRenewableResource,InstituteforAdvancedLearningandResearch,150SlaytonAve,Danville,VA24540,USA.
2DepartmentofHorticulture,VirginiaPolytechnicInstituteandStateUniversity,Blacksburg,VA24601,USA.
3DepartmentofForestResourcesandEnvironmentalConservation,VirginiaPolytechnicInstituteandStateUniversity,Blacksburg,VA24601,USA.
4TheDewelMicroscopyFacilityattheCollegeofArtsandSciences,AppalachianStateUniversity,Boone,NC28608,USA.
Received:26March2012Accepted:10May2012Published:30May2012References1.
HamelinckCN,HooijdonkDV,FaaijAP:Ethanolfromlignocellulosicbiomass:techno-economicperformanceinshort-,middle-,andlong-term.
BiomassBioenergy2005,28:384–410.
2.
ErahinME,GomecCY,DereliRK,ArikanO,OzturkI:Biomethaneproductionasanalternativebioenergysourcefromcodigesterstreatingmunicipalsludgeandorganicfractionofmunicipalsolidwastes.
JBiomedBiotechnol2011,201(1):951043.
doi:10.
1155/2011/951043.
3.
SissineF:EnergyIndependenceandSecurityAct.
;2007[http://www.
energy.
senate.
gov/public/index.
cfm/files/serveFile_id=82061c68-eef5-4fc3-81b6-ecd569a1d8a3].
4.
NationalBiofuelsActionPlan:2008[http://www1.
eere.
energy.
gov/biomass/pdfs/nbap.
pdf].
5.
SandersonM,AdlerPR,BoatengAA,CaslerMD,SarathG:SwitchgrassasabiofuelsfeedstockintheUSA.
CanJPlantSci2006,86:1315–1325.
6.
McLaughlinSB,KszosLA:Developmentofswitchgrass(Panicumvirgatum)asabioenergyfeedstockintheUnitedStates.
BiomassBioenergy2005,28:515–535.
7.
HillJ,NelsonE,TilmanD,PolaskyS,TiffanyD:Environmental,economic,andenergeticcostsandbenefitsofbiodieselandethanolbiofuels.
ProcNatlAcadSciUSA2006,103:11206–11210.
8.
HumphreysMO:Thecontributionofconventionalplantbreedingtoforagecropimprovement.
Saskatoon:Proceedingsofthe18thInternationalGrasslandCongress;1999.
9.
LiebigMA,JohnsonHA,HansonJD,FrankAB:Soilcarbonunderswitchgrassstandsandcultivatedcropland.
BiomassBioenergy2005,28:347–354.
10.
McLaughlinSB,UgarteDGLT,GartenCT,LyndLR,SandersonMA,TolbertVR,WolfDD:High-valuerenewableenergyfromprairiegrasses.
EnvironSciTechnol2002,36:2122–2129.
11.
DyerJM,MullenRT:Engineeringplantoilsashigh-valueindustrialfeedstocksforbiorefining:theneedforunderpinningcellbiologyresearch.
PhysiolPlant2008,132:11–22.
12.
SturzAV,ChristieB,NowakJ:Bacterialendophytes:potentialroleindevelopingsustainablesystemofcropproduction.
CritRevPlantSci2000,19:1–30.
13.
BergG:Plant-microinteractionspromotingplantgrowthandhealth:perspectivesforcontrolleduseofmicroorganismsinagriculture.
ApplMicrobialBiotechnol2009,84:11–18.
14.
MeiC,FlinnBS:Theuseofbeneficialmicrobialendophytesforplantbiomassandstresstoleranceimprovement.
RecentPatentsonBiotechnol2010,4:81–95.
15.
FunkCR,WhiteRH,BreenJP:ImportanceofAcremoniumendophytesinturf-grassbreedingandmanagement.
AgricEcosystEnviron1993,44:215–232.
16.
GhimireSR,CharltonND,CravenKD:Themycorrhizalfungus,sebacinavermifera,enhancesseedgerminationandbiomassproductioninswitchgrass(PanicumvirgatumL).
BioenergyRes2009,2:51–58.
17.
GhimireSR,CharltonND,BellJD,KrishnamurthyYL,CravenKD:Biodiversityoffungalendophytecommunitiesinhabitingswitchgrass(PanicumvirgatumL)growinginthenativetallgrassprairieofnorthernOklahoma.
FungalDiversity2010,47:19–27.
18.
KhanSA,HamayunM,YoonH,KimHY,SuhSK,HwangSK,KimJM,LeeIJ,ChooYS,KongWS,LeeBM,KimJG:PlantgrowthpromotionandPenicilliumcitrinum.
BMCMicrobial2008,8:231–240.
19.
NowakJ,MeiC,LowmanS,ZhaoB,SeilerJ,FlinnB:Developmentofswitchgrass(PanicumvirgatumL.
)formarginallandsbasedongenotypiccompatibilitywithbeneficialbacteria[abstract].
Dalian:InFirstAnnualWorldCongressofBioenergy;2011:25–29.
20.
WelbaumG,SturzAV,DongZ,NowakJ:Managingsoilmicroorganismstoimproveproductivityofagroecosystems.
CritRevPlantSci2004,23:175–193.
21.
CompantS,DuffyB,NowakJ,ClémentC,BarkaEA:Useofplantgrowth-promotingbacteriaforbiocontrolofplantdiseases:principles,mechanismsofactionandfutureprospects.
ApplEnvironMicrobiol2005,71:4951–4959.
22.
CompantS,ReciterB,SessitschA,NowakJ,ClementC,BarkaEA:EndophyticColonizationofVitisvinifereL.
byPlantGrowth-PromotingBacteriumBurkholderiasp.
StrainPsJN.
ApplEnvironMicrobiol2005,71:1685–1693.
23.
GlickBR,PenroseDM,LiJ:Amodelfortheloweringofplantethyleneconcentrationsbyplantgrowthpromotingbacteria.
JTheorBiol1998,190:63–68.
24.
GlickBR:BacterialACCdeaminaseandthealleviationofplantstress.
AdvApplMicrobiol2004,56:291–312.
25.
MattosKA,PáduaVLM,RomeiroA,HallackLF,NevesBC,UlissesTMU,BarrosCF,TodeschiniAR,PreviatoJO,Mendona-PreviatoL:EndophyticcolonizationofricebythedizaotrophicbacteriumBurkholderiakururiensisanditsabilitytoenhanceplantgrowth.
AnAcadBrasCienc2008,80:477–493.
26.
ArshadM,FrankenbergerWT:Microbialproductionofplanthormones.
PlantSoil1991,133:1–8.
Kimetal.
BiotechnologyforBiofuels2012,5:37Page9of10http://www.
biotechnologyforbiofuels.
com/content/5/1/3727.
LazarovitsG,NowakJ:Rhizobacteriaforimprovementofplantgrowthandestablishment.
Hortscience1996,32:188–192.
28.
ConnKL,NowakJ,LazarovitsG:Agnotobioticbioassayforstudyinginteractionsbetweenpotatoesandplantgrowth-promotingrhizobacteria.
CanJMicrobiol1997,43:801–808.
29.
BarkaEA,BelarbiA,HachetC,NowakJ,AudranJC:EnhancementofinvitrogrowthandresistancetograymouldofVitisviniferaco-culturedwithplantgrowth-promotingrhizobacteria.
FEMSMicrobiolLett2000,186:91–95.
30.
NowakJ,AsieduSK,LazarovitsG:Enhancementofinvitrogrowthandtransplantstresstoleranceofpotatoandvegetableplantsco-culturedwithaplantgrowthpromotingrhizobacterium.
InEcophysiologyandPhotosyntheticInVitroCultures.
EditedbyCarreF,ChagvardieffP.
Aix-en-Provence:CEA;2003:173–180.
31.
SessitschA,CoenyeT,SturzAV,VandammeP,BarkaEA,SallesJF,ElsasJD,FaureD,ReiterB,GlickBR,Wang-PruskiG,NowakJ:Burkholderiaphytofirmanssp.
nov.
,anovelplant-associatedbacteriumwithplant-beneficialproperties.
IntJSystEvolMicrobiol2005,55:1187–1192.
32.
WangK,ConnK,LazarovitsG:InvolvementofquinolinatephophoribosyltransferaseinpromotionofpotatogrowthbyaBurkholderiaStrain.
ApplEnvironMicrobiol2006,72:760–768.
33.
WeilharterA,MitterB,ShinMV,ChainPSG,NowakJ,SessitschA:Completegenomesequenceoftheplantgrowth-promotingendophyteBurkholderiaphytofirmansstrainPsJN.
JBacteriol2011,193:3383–3384.
34.
BarkaEA,NowakJ,ClémentS:Enhancementofchillingresistanceofinoculatedgrapevineplantletswithaplantgrowth-promotingrhizobacterium,BurkholderiaphytofirmansstrainPsJN.
ApplEnvironMicrobiol2006,72:7246–7252.
35.
BensalimS,NowakJ,AsieduSK:Aplantgrowthpromotingrhizobacteriumandtemperatureeffectsonperformanceof18clonesofpotato.
AmJPotatoRes1998,75:145–152.
36.
FrommelMI,NowakJ,LazarovitsG:Growthenhancementanddevelopmentalmodificationofinvitrogrownpotato(Solanumtuberosumspp.
tuberosum)asaffectedbyanonfluorescentPseudomonassp.
PlantPhysiol1991,96:928–936.
37.
SharmaV,NowakJ:Enhancementofverticilliumwiltresistanceintomatotransplantsbyinvitroco-cultureofseedlingswithaplantgrowthpromotingrhizobacterium(Pseudomonassp.
strainPsJN).
CanJMicrobiol1998,44:528–536.
38.
BarkaEA,GogniesS,NowakJ,AudranJ,BelarbiA:InhibitoryeffectofendophytebacteriaonBotrytiscinereaanditsinfluencetopromotegrapevinegrowth.
BiolControl2002,24:135–142.
39.
ReiterB,PfeiferU,SchwabH,SessitschA:ResponseofendophyticbacterialcommunitiesinpotatoplantstoinfectionwithErwiniacaratovorasubsp.
atroseptica.
ApplEnvironMicrobiol2002,68:2261–2268.
40.
WhippsJM:Microbialinteractionsandbiocontrolintherhizosphere.
JExpBot2001,52:487–511.
41.
LongHH,SchmidtDD,BaldwinIT:Nativebacterialendophytespromotehostgrowthinaspecies-specificmanner;phytohormonemanipulationsdonotresultincommongrowthresponse.
PLoSOne2008,3:e2702.
42.
PenroseDM,GlickBR:MethodsforisolatingandcharacterizingACCdeaminase-containingplantgrowthpromotingrhizobacteria.
PhysiolPlant2003,118:10–15.
43.
NowakJ,AsieduSK,BensalimS,RichardsJ,StewartA,SmithC,StevensD,SturzAV:Fromlaboratorytoapplications:challengesandprogresswithinvitrodualculturesofpotatoandbeneficialbacteria.
PlantCellTissOrgCult1998,52:97–103.
44.
NowakJ,VeilleuxRE,NowakJ,TurgeonS:Primingfortransplantstressresistanceininvitropropagationviaplantletbacterization.
ActaHort2007,748:65–75.
45.
PillayVK,NowakJ:Inoculumdensity,temperature,andgenotypeeffectsoninvitrogrowthpromotionandepiphyticandendophyticcolonizationoftomato(LycopersiconesculentumL.
)seedlingsinoculatedwithapseudomonadbacterium.
CanJMicrobiol1997,43:354–361.
46.
MurashigeT,SkoogF:Arevisedmediumforrapidgrowthandbioassayswithtobaccotissuecultures.
PhysiolPlant1962,15:473–497.
doi:10.
1186/1754-6834-5-37Citethisarticleas:Kimetal.
:Growthpromotionandcolonizationofswitchgrass(Panicumvirgatum)cv.
AlamobybacterialendophyteBurkholderiaphytofirmansstrainPsJN.
BiotechnologyforBiofuels20125:37.
SubmityournextmanuscripttoBioMedCentralandtakefulladvantageof:ConvenientonlinesubmissionThoroughpeerreviewNospaceconstraintsorcolorgurechargesImmediatepublicationonacceptanceInclusioninPubMed,CAS,ScopusandGoogleScholarResearchwhichisfreelyavailableforredistributionSubmityourmanuscriptatwww.
biomedcentral.
com/submitKimetal.
BiotechnologyforBiofuels2012,5:37Page10of10http://www.
biotechnologyforbiofuels.
com/content/5/1/37
- Average515hh相关文档
- ryngeal515hh
- www.scholarsresearchlibrary.com
- conven515hh
- Actively515hh
- 1.3515hh
- rewrite515hh
ucloud国内云服务器2元/月起;香港云服务器4元/首月;台湾云服务器3元/首月
ucloud云服务器怎么样?ucloud为了扩大云服务器市场份额,给出了超低价云服务器的促销活动,活动仍然是此前的Ucloud全球大促活动页面。目前,ucloud国内云服务器2元/月起;香港云服务器4元/首月;台湾云服务器3元/首月。相当于2-4元就可以试用国内、中国香港、中国台湾这三个地域的云服务器1个月了。ucloud全球大促仅限新用户,国内云服务器个人用户低至56元/年起,香港云服务器也仅8...
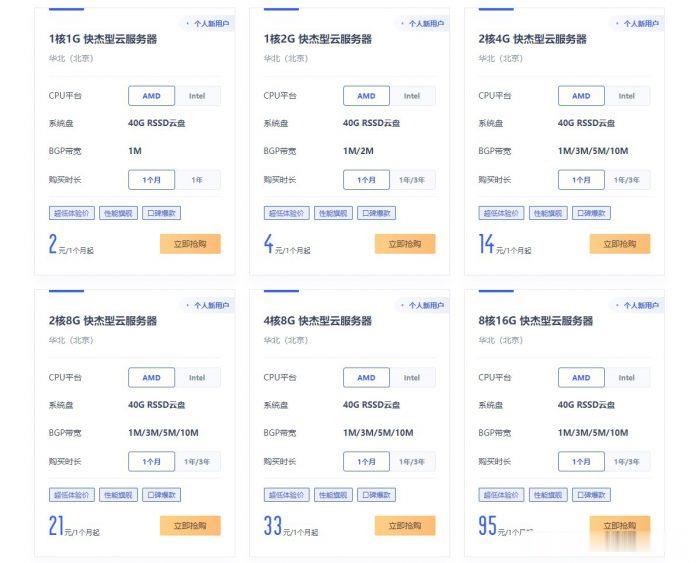
DogYun(300元/月),韩国独立服务器,E5/SSD+NVMe
DogYun(中文名称狗云)新上了一批韩国自动化上架独立服务器,使用月减200元优惠码后仅需每月300元,双E5 CPU,SSD+NVMe高性能硬盘,支持安装Linux或者Windows操作系统,下单自动化上架。这是一家成立于2019年的国人主机商,提供VPS和独立服务器租用等产品,数据中心包括中国香港、美国洛杉矶、日本、韩国、德国、荷兰等。下面分享这款自动化上架韩国独立服务器的配置和优惠码信息。...

Boomer.Host(年付3.5美)休斯敦便宜VPS
Boomer.Host是一家比较新的国外主机商,虽然LEB自述 we’re now more than 2 year old,商家提供虚拟主机和VPS,其中VPS主机基于OpenVZ架构,数据中心为美国得克萨斯州休斯敦。目前,商家在LET发了两款特别促销套餐,年付最低3.5美元起,特别提醒:低价低配,且必须年付,请务必自行斟酌确定需求再入手。下面列出几款促销套餐的配置信息。CPU:1core内存:...
515hh com为你推荐
-
苹果5怎么越狱苹果5怎么越狱安装迅雷看看播放器迅雷看看播放器下了安装不了idc前线钢铁雄心2修改器2012年正月十五农历2012年正月15早上9点多生的!命里缺什么!是什么命相cr2格式如何打开CR2格式的照片呢系统分析员系统分析师是什么职业?主要做什么工作?怎么上传音乐怎么上传音乐到网上blogcn远目是什么意思?QzongQZONG怎么玩?微信怎么看聊天记录微信在手机上怎么查看聊天记录